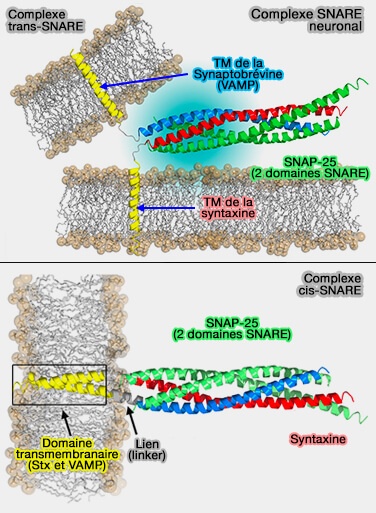
Le complexe SNARE est aussi appelé SNAREpin en anglais.
Complexe SNARE neuronal
(Figure : vetopsy.fr d'après Fang et Lindau)
Lors de la fusion, les complexes trans-SNARE passent en position cis, i.e. complexes cis-SNARE dans lesquels toutes les protéines SNARE sont positionnées sur la même membrane.
Rôles du lien et du domaine transmembranaire
1. Le complexe cis-SNARE neuronal, qui représenterait l'état post-fusion, montre que non seulement les domaines SNARE, mais aussi les domaines transmembranaires (TMD) peuvent s'associer.
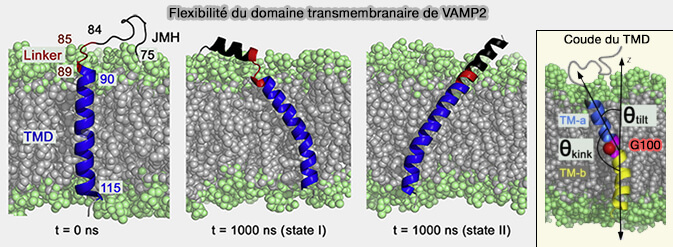
Pour accéder à cette force motrice potentielle déjà pendant l'hémifusion, les domaines transmembranaires (TMD) doivent posséder une flexibilité mécanique suffisante pour permettre leur association.
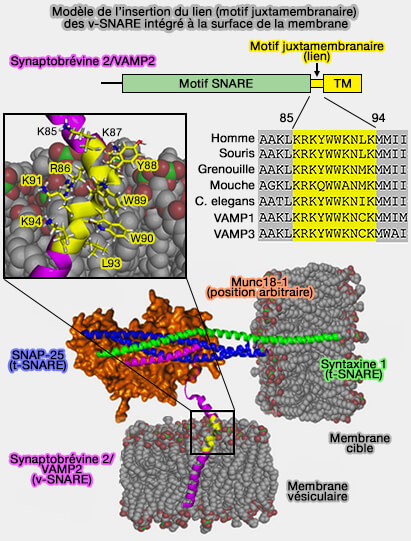
aux résidus non polaires du motif juxtamembranaire de s'insérer dans la phase hydrophobe de la bicouche,
aux résidus basiques de s'intégrer dans la phase hydrophile de la membrane.
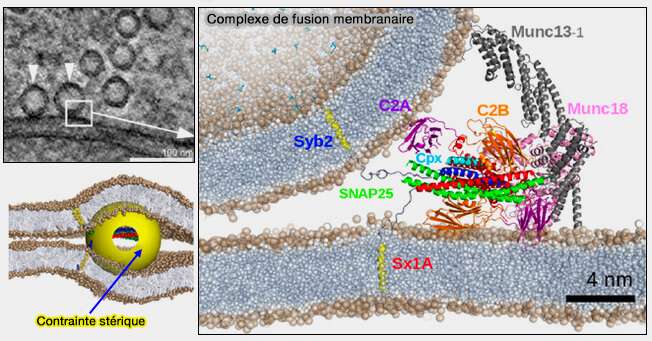
Sur la figure, les atomes de carbone, d'oxygène et de phosphore de la bicouche lipidique sont respectivement colorés en gris, rouge et vert. Étant donné que la structure de Munc18-1 liée au complexe trans-SNARE à demi-zippé n'a pas été déterminée, la position de Munc18-1 représentée dans le modèle est arbitraire.
et/ou de résidus β-ramifiés isoleucine/valine, dirigés éventuellement via l'interaction entre Lys94 et du motif juxtamembranaire et les lipides (courbure ou bending).
Flexibilité du domaine transmembranaire (TMD) de la synaptobrévine 2/VAMP2 (syb2)
(Figure : vetopsy.fr d'après Han et coll)
Cette flexibilité conformationnelle faciliterait la transition de l'hémifusion à la fusion qui implique la conversion du complexe trans-SNARE en complexe cis-SNARE.
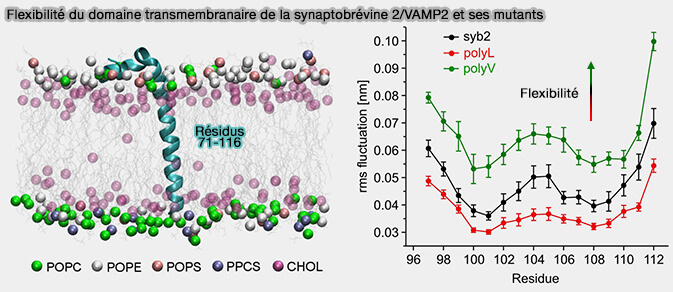
L'introduction de résidus de leucine stabilisant l'hélice du domaine transmembranaire (TMD) recouvrant le feuillet externe de la vésicule altère fortement l'exocytose et ralentit la fusion et la dilatation des pores.
En revanche, l'augmentation du nombre de valine déstabilisant l'hélice à ramification β ou les résidus d'isoleucine dans le TMD, restaurent la sécrétion normale, mais accélèrent l'expansion des pores de fusion au-delà du taux trouvé pour la protéine de type sauvage.
Flexibilité du domaine transmembranaire (TMD) de la synaptobrévine 2/VAMP2 (syb2)
(Figure : vetopsy.fr d'après Dhara et coll)
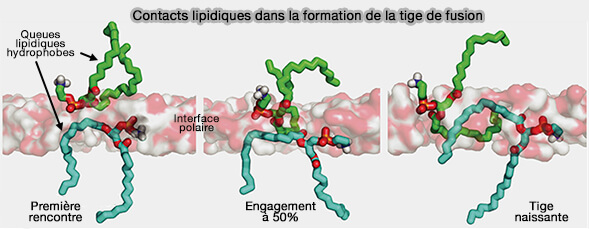
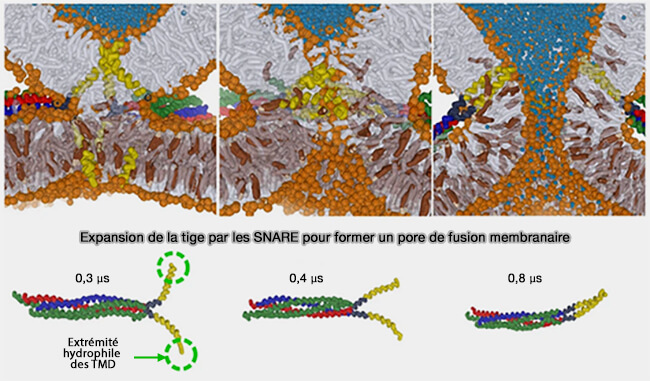
L'état de transition pour la formation de la tige se produit lorsqu'une paire de queues lipidiques entre en contact à travers la couche d'interface polaire. Dans les simulations, le contact entre une seule paire de queues hydrophobes serait suffisant pour nucléer la formation de tiges de fusion ( rôles des lipides dans la formation de la tige).
Contacts lipidiques dans la formation de la tige de fusion
(Figure : vetopsy.fr d'après Kasson et coll)
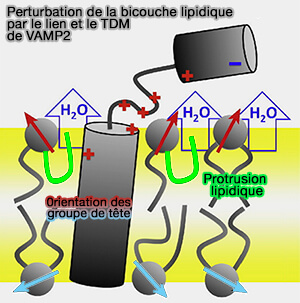
Perturbations lipidique de VAMP2
(Figure : vetopsy.fr d'après Han et coll)
influence l'orientation du groupe de tête (flèches rouges et jaunes).
La déshydratation de la membrane et l'hydrophobicité de la surface de la membrane réduisent ainsi les forces répulsives entre les deux membranes qui se rapprochent.
L'oligomérisation des SNARE augmente probablement encore la protrusion lipidique, probablement aussi en induisant une courbure de la membrane.
Ces deux phénomènes abaisseront la barrière énergétique pour la formation des pores de la membrane.
Les modifications de la structure membranaire induites par les SNARE seraient plus le fait du lien et de JM et, dans une moindre mesure, du TDM par la perturbation de la bicouche lipidique.





 rôles des lipides dans la formation de la tige).
rôles des lipides dans la formation de la tige).