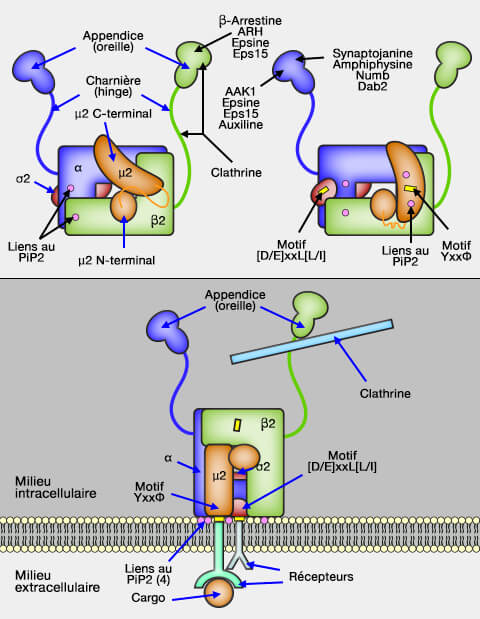
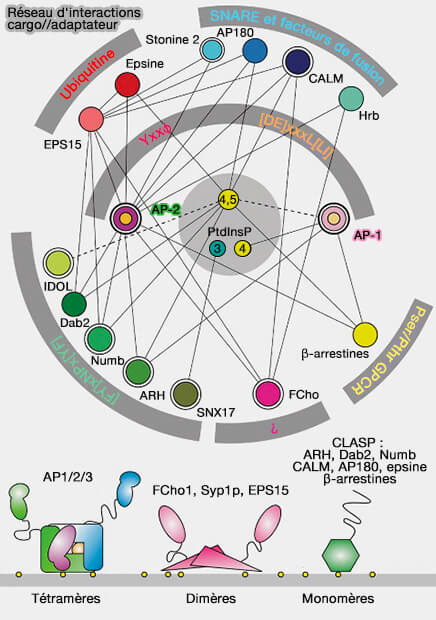
Elles engagent et accumulent des cargos sur une membrane source en se liant aux signaux de tri situés dans les domaines cytoplasmiques du cargo.
Elles recrutent une couche externe et/ou d'autres protéines accessoires pour générer des transporteurs qui facilitent la livraison du cargo à sa destination.
La protéine adaptatrice AP-2 est un hétérotétramère constitué de sous-unités α, β2, µ2 et σ2 qui fonctionne comme un adaptateur de la clathrine.
Réseau d'interactions cargo/adaptateurs
(Figure : vetopsy.fr d'après Traub et Bonifacino)
AP-2 recrute les cargos, via leurs signaux peptidiques, vers les vésicules.
1. Deux grandes sous-unités (α et β2 de 100 kDa) sont formées par :
un domaine N-terminal qui se lie avec la moyenne (μ2) et la petite sous-unité (σ2) pour former le corps de l'adaptine,
un domaine moyen ayant un rôle de charnière (hinge),
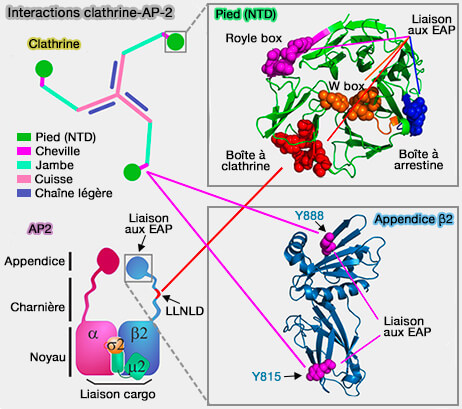
un domaine C-terminal, l'appendice ou l'oreille, relié au noyau par l'intermédiaire d'un long segment flexible qui interagit avec la clathrine, diverses adaptines ou des protéines accessoires (cf. figure).
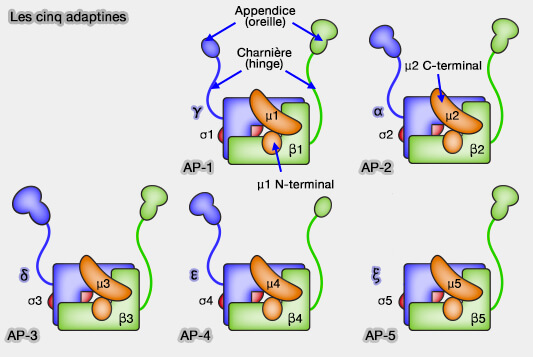
Remarque : n'oubliez pas que la sous-unité α de l'AP-2 est appelée γ//δ/ε/ζ pour les AP-1, AP-3, AP-4 et AP-5.
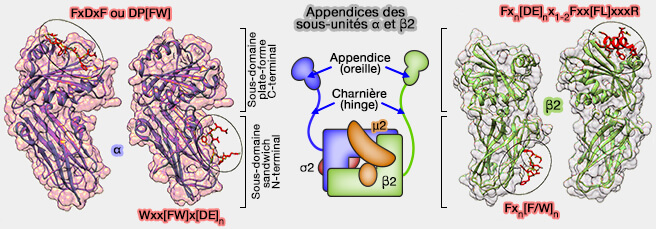
2. Les appendices des sous-unités α et β2 d'AP-2 partagent une structure bilobée similaire, chacun consistant en un sous-domaine dit sandwich N-terminal attaché à un sous-domaine dit plate-forme C-terminal.
Chaque sous-domaine de chaque appendice contient une surface d'interaction distincte pour la liaison au partenaire protéique, i.e. une seule molécule AP-2 possède 4 sites de liaison séparés.
La spécificité pour chaque site est conférée par des motifs d'interaction courts qui sont caractérisés par des chaînes latérales aromatiques.
Interactions clathrine-AP-2
(Figure : vetopsy.fr d'après Chen et coll)
Remarque : le noyau (ou coeur) est formé par toutes les sous-unités, excepté les appendices.
Interactions de α
1. La sous-unité α interagit avec les motifs DP[F/W], FxDxF et Wxx[FW]x[DE] et [D/E]xxL[L/I] du cargo.
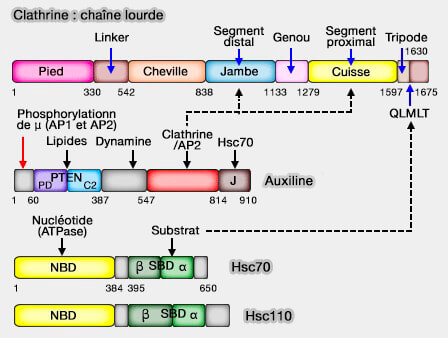
à la clathrine (domaine C-terminal de la chaîne lourde) par 2 séquences LΦxΦD/E (boîtes à clathrine ou CBM : Clathrin-Binding Motif) chez les mammifères (leucine, Φ - acide aminé hydrophobe - et x - acide aminé polaire - D/E en dernier) qui sont situées une sur sa charnière et une autre sur son oreille,
Plus généralement, les protéines CLASP élargissent le répertoire de cargos endocytaires triés dans des vésicules recouvertes de clathrine au-delà des protéines transmembranaires qui se lient physiquement à l'adaptateur AP-2.
Motifs des appendices α2 et β2
(Figure : vetopsy.fr d'après Smith et coll)
2. Les LDLR et les GPCR sont internalisés par l'ARH et les β-arrestines, respectivement, qui se lient à la plate-forme de l'appendice AP-2, via un motif α-hélicoïdal [D/E]x1-2Fxx[F/L]xxxR, motif également présent et fonctionnel dans les epsines ( internalisation des GPCR).
Sous-unité moyenne : μ2
Structure
Recrutement de l'AP-2
(Figure : vetopsy.fr)
1. La sous-unité moyenne (μ2 de 47-50 kDa) possède :
un domaine N-terminal qui contribue au corps de l'adaptine avec α, β2 et σ2,
un domaine C-terminal qui reconnait le peptide signal du récepteur.
2. Le peptide signal est une séquence particulière du domaine cytosolique du récepteur qui se lie au domaine C-terminal de la sous-unité μ pour l'intégration dans le puits de clathrine et peut différer selon les cas.
YxxΦ, où l'acide aminé x polaire est spécifique pour l'adaptine et Φ est un acide aminé hydrophobe.
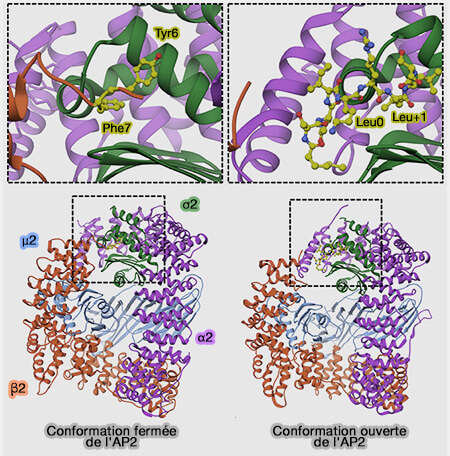
3. Toutefois, l'interaction entre les sous-unités C-terminale de μ2 et β2 (conformation fermée) masque le motif YxxΦ de reconnaissance du récepteur et AP-2 montre une faible affinité pour le cargo et la membrane.
Elle se fixe sur l'oreille de la sous-unité α, ce qui multiplie par 100 son affinité pour le cargo et la membrane.
Cette phosphorylation provoque un changement conformationnel de μ2 (rotation de 129° autour de son grand axe et translation de 39 Å, i.e. la moitié de sa longueur, par rapport à son domaine N-terminal) qui expose le motif de liaison YxxΦ.
Ce domaine se lie aussi à 2 PI(4,5)P2 ou PIP2 pour laisser le motif exposé.
Cette phosphorylation est très fortement stimulée par la clathrine.
2. L'auxiline/GAK, protéines homologues, i.e. cyclin G–Associated kinase ubiquiste et auxiline purement cérébrale, peut aussi intervenir dans le processus d'endocytose chez les mammifères, bien qu'elle ait aussi d'autres rôles.
L'auxiline/GAK est étudiée dans un chapitre spécial.
Le réaménagement structurel de l'AP-2 rend coplanaires les 4 liaisons PIP2 [phosphatidylinositol (4,5)-bisphosphate ou PtdIns(4,5)P2] et les deux motifs de liaison endocytaires pour faciliter les interactions avec la membrane et le cargo.





 internalisation des GPCR).
internalisation des GPCR).