de tube, de pont et de tunnel multimériques ou monomériques, ce qui a l'énorme avantage de déplacer le lipide dit par glissement (sliding en anglais) sans ou avec un minimum de mouvements des protéines, i.e. ce qui permet une augmentation substantielle de la vitesse du transfert.
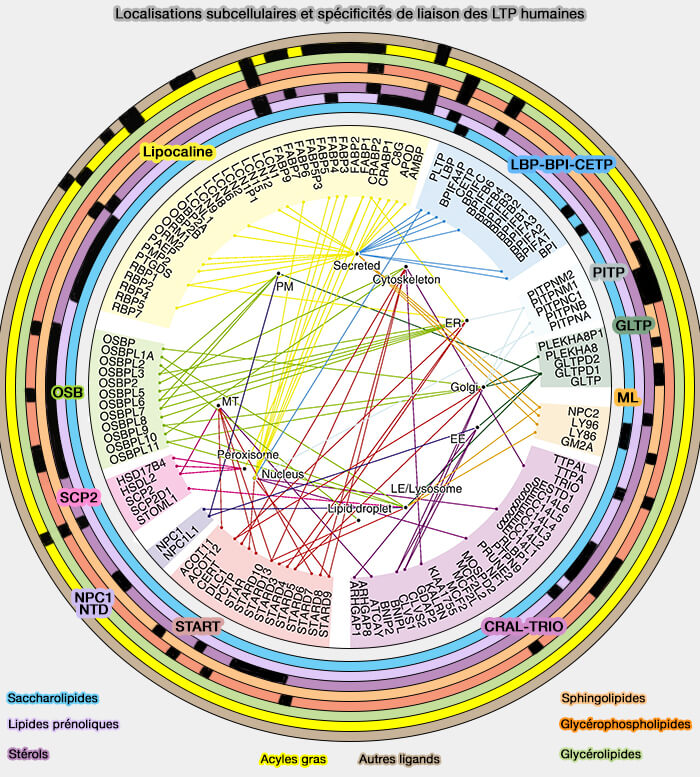
Localisations subcellulaires et spécificités de liaison des LTP humaines
(Figure : vetopsy.fr d'après Chiapparino et coll)
Vue d'ensemble des transferts lipidiques par LTP tubulaires
1. Les LTP sont de forme tubulaire avec un tunnel hydrophobe apparemment adapté à la canalisation des lipides d'une extrémité à l'autre par glissement lipidique.
Les domaines SMP de la famille TULIP semble posséder un tunnel hydrophobe apparemment adapté à la canalisation des lipides d'une extrémité à l'autre par glissement lipidique.
On peut aussi ranger dans cette catégorie le complexe Lpt des bactéries.
2. En raison de la pénalité énergétique associée à l'enfouissement des groupes de tête polaires dans une cavité hydrophobe, certaines de ces LTP tubulaires présentent une ouverture latérale appelée " couture " (seam en anglais) qui longe le tube/barillet et dont les bords sont parfois délimités par des hélices α torsadées.
Cette couture accessible aux solvants accueille les groupe de tête hydrophiles tandis que les sa chaîne acyle (R-C(=O)-) restent enfouies dans le noyau de la LTP et permet le glissement des lipides le long du noyau tubulaire.
Remarque : selon les auteurs, ces LTP peuvent aussi être classées dans les des ponts car elles joignent les deux membranes.
3. Alors que les navettes en forme de boîte ont un point d'accès unique à leurs cavités de liaison, les LTP tubulaires forment un long pont hydrophobe qui s'ouvre de chaque côté de la membrane et qui permet aux queues lipidiques hydrophobes de coulisser au travers.
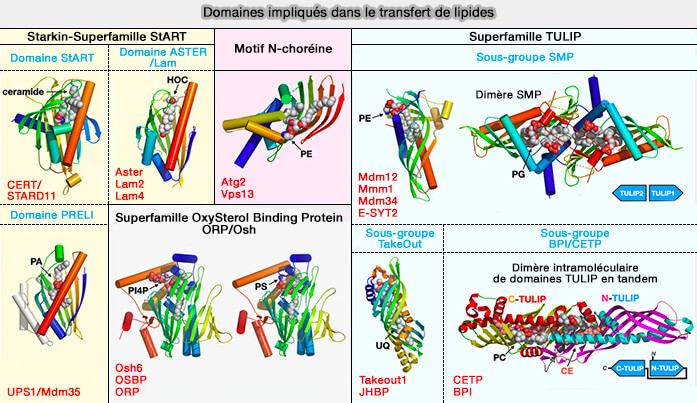
Domaines impliqués dans le transfert des lipides
(Figure : vetopsy.fr d'après Egea)
4. Nous prendrons comme exemples :
a. le complexe LTP de la levure mutimérique qui pourrait aussi être classé dans les ponts.
b. des protéines TULIP à domaine SMP :
le complexe ERMES de la levure multimérique,
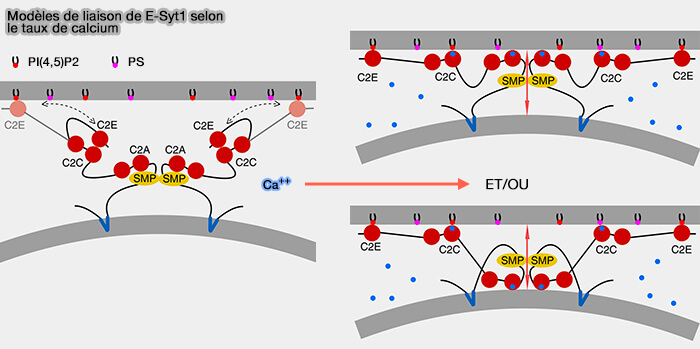
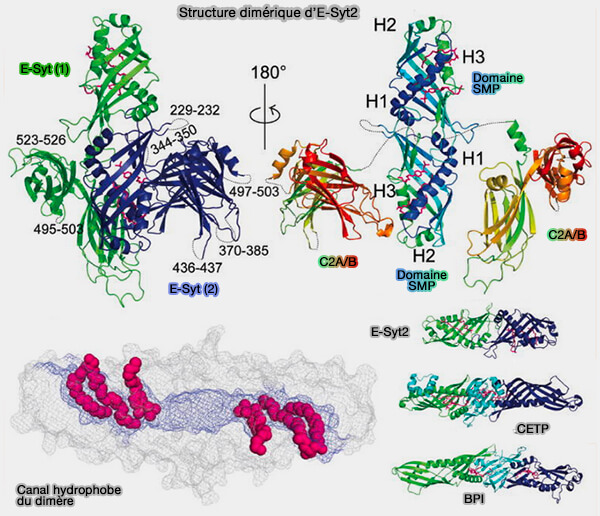
les E-SYST des vertébrés qui se dimérisent,
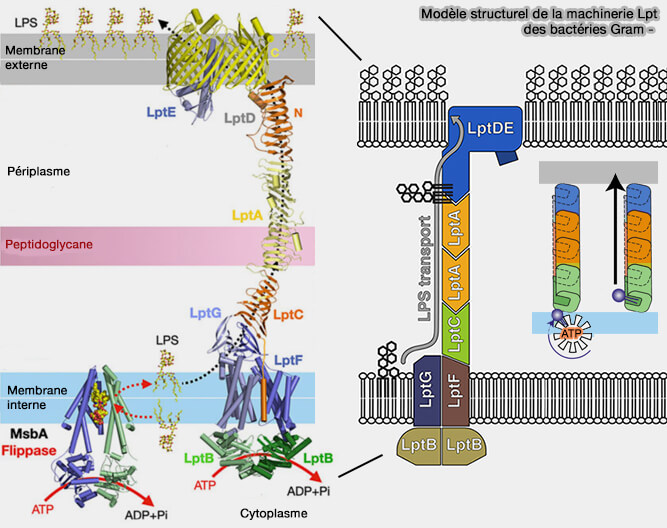
Transport du lipopolysaccharide (Lpt) des bactéries gram -
Les sept protéines de ce système (LptABCDEFG) forment deux sous-complexes, i.e. LptBFGC dans la membrane interne et LptDE dans la membrane externe, reliés par LptA.
Le LPS est transporté à travers la membrane externe et inséré dans le feuillet externe par le complexe LptDE.
LPT tubulaires des eucaryotes (TULIP)
Les TULIP (TUbular LIPid binding proteins ou protéines tubulaires de liaison aux lipides) forment des tunnels hydrophobes pour la canalisation des lipides d'une extrémité à l'autre par glissement lipidique.
Ces TULIP contiennent des domaines SMP (Synaptotagmin-like Mitochondrial lipid-binding Protein) qui sont exclusivement associés aux sites de contact membranaire (MCS).
a. Les résidus à l'interface des deux domaines SMP sont parmi les plus conservés, ce qui correspond à la pertinence physiologique du dimère, et ils interviennent très probablement dans l'homo- et l'hétérodimérisation des E-Syt.
Il se connecte au solvant aux deux extrémités et via une couture étroite sur toute la longueur du cylindre, comme dans le cas du sytème lpt des bactéries.
Remarque : on ne trouve pas de similarité de séquence primaire significative entre le domaine SMP des E-Syt et les protéines de repli connu.
b. Toutefois, BPI et CETP ne se dimérisent pas, car chaque monomère comprend deux domaines de type SMP plutôt qu'un, les deux modules étant disposés en tandem et reliés par une feuille bêta non présente dans les E-Syt ( cf. figure au-dessus).
1. Les phospholipides (PL) peuvent se déplacer par un mécanisme de « glissement » (sliding en anglais) le long de la couture (seam) latérale du domaine SMP avec leur tête polaire exposée au solvant tandis que les chaînes acyle restent enfouies dans le domaine SMP.
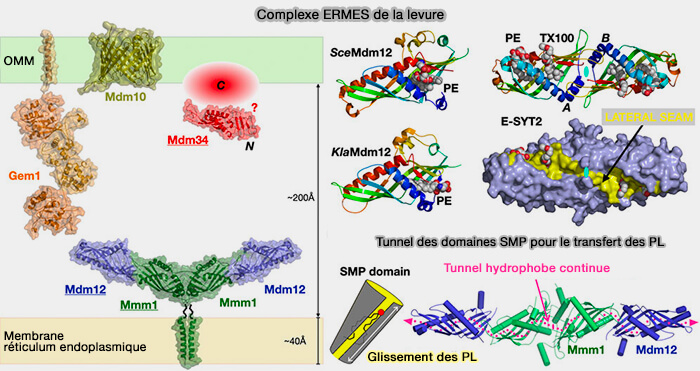
Complexe ERMES de la levure
(Figure : vetopsy.fr d'après Egea)
Les mutations introduites à l'interface Mmm1-à-Mdm12 bloquent l'échange lipidique in vitro.
b. Les courbures observées dans le dimère E-SYT2 et le tétramère Mdm12-Mmm1 pourraient être importantes lors de l'étape de désorption lipidique pour moduler la forme de la membrane et inversement (étapes limitante du transfert lipidique).
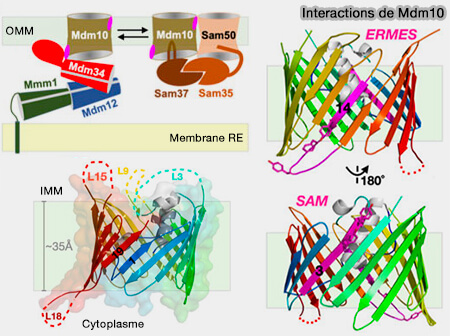
2. Dans le complexe ERMES, si le rôle de Mdm12/Mmm1 est acquis, le rôle de Mdm34 reste obscur.
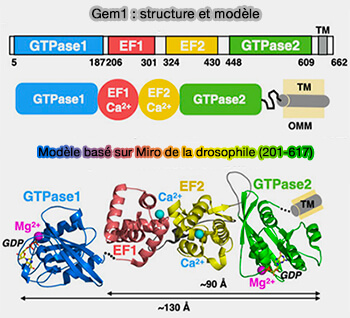
Gem1 : structure et modèle
(Figure : vetopsy.fr d'après Egea)
a. Cette GTPase régulatrice de liaison au calcium est une protéine TA (Tail-Anchored, i.e. ancrée par la queue), i.e. à une seule hélice transmembranaire C-terminale.
Sur ces bases, plusieurs modèles ont été proposés, basés sur la forme tubulaire du domaine SMP et la présence d'un long canal hydrophobe continu s'étendant sur environ 190 Å dans le tétramère Mdm12-Mmm1.
Ce canal serait apparemment adapté et suffisant pour relier les deux membranes apposées et transférer les lipides d'une membrane à l'autre.
Ces modèles ne prennent cependant pas en compte le rôle de Mdm34, la dernière protéine contenant un domaine SMP.
Ce modèle combine un " glissement lipidique " dans Mdm12/Mmm1 et une étape de transfert vers Mdm34.
Les domaines SMP fonctionnent non seulement comme des LTP, mais également comme des échafaudages polyvalents pour établir des interfaces protéine-protéine spécifiques.
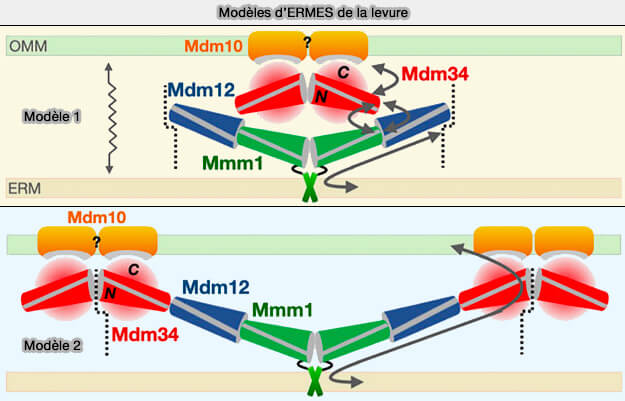
Modèles d'ERMES de la levure
(Figure : vetopsy.fr d'après Egea)
2. Un deuxième modèle inclut Mdm34 à Mmm1 et Mdm12 pour former un ensemble tubulaire plus long, formant un tunnel quasi continu reliant les deux organites.
L'interaction entre Mdm12 et Mdm34 serait plus faible, conférant ainsi la flexibilité et la fugacité nécessaires à l'assemblage ternaire.
Quel que soit le modèle, ces mécanismes semblent bien adaptés pour soutenir le transfert " en masse " des lipides nécessaires à l’alimentation des structures membranaires dynamiques telles que les mitochondries.





 cf. figure au-dessus).
cf. figure au-dessus).