


Système endo-lysosomal
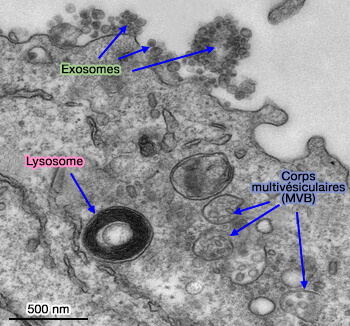
Exosomes
Biogenèse et libération
- Vue d'ensemble du système endomembranaire
- Endosomes
- Vue d'ensemble des endosomes
- Endosomes précoces
- Endosomes de recyclage
- Vésicules intraluminales (ILV) et endosomes ou corps mutivésiculaires (MVE/MVB)
- Endosomes tardifs
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La biogenèse et la libération des exosomes peuvent s'effectuer par plusieurs voies.
Biogenèse des exosomes
La biogenèse des exosomes comprend plusieurs étapes et bon nombre des composants sont également impliqués dans d'autres voies du trafic vésiculaire (Biogenesis and secretion of exosomes 2014 et New insights into extracellular vesicle biogenesis and function 2019).
(Figure : vetopsy.fr d'après Edgar)
Ce processus rappelle l'exocytose régulée des organites liés aux lysosomes (LRO) dans des types de cellules spécialisées (The Biogenesis of Lysosomes and Lysosome-Related Organelles 2014).
Les sous-populations de MVE/MVB utilisent des machines différentes pour la biogenèse des exosomes dans différents types de cellules et/ou qui coexistent dans le même type cellulaire (Signaling Pathways in Exosomes Biogenesis, Secretion and Fate 2013).
Rôle de la formation des ILV
La formation des vésicules intraluminales (ILV) peut être considérée comme la première étape du processus.
De manière constante, la biogenèse des exosomes dépend (Q&A: What are exosomes, exactly? 2019) :
- soit du complexe ESCRT, de manière dépendante de l'ubiquitination ou non (Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles 2013),
- soit indépendamment de ce complexe, utilisant le plus souvent des tétraspanines,
- soit d'ALIX (Syndecan–syntenin–ALIX regulates the biogenesis of exosomes 2012 et Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2 2013).
1. La déplétion des composants du complexe ESCRT affecte la taille de l'exosome, la quantité et la cargaison protéique (Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles 2013).
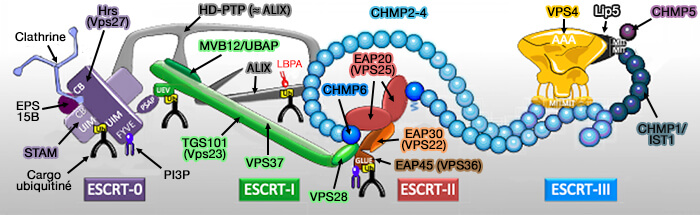
(Figure : vetopsy.fr d'après Christ et Hill)
Le complexe ESCRT est composé de quatre complexes protéiques (ESCRT-0, I, II et III) et, avec les protéines accessoires (ALIX, Vps4) associées à la membrane endosomale de manière coordonnée, régule la sélection du cargo et la formation des ILV ( formation des ILV par la voie dépendante de ESCRT).
formation des ILV par la voie dépendante de ESCRT).
- ESCRT 0 est recruté à la membrane endosomale par des protéines transmembranaires monoubiquitées.
- ESCRT I et II promeuvent l'invagination de la membrane contenant ces complexes.
- ESCRT III termine la scission de la membrane, générant des ILV (Signaling Pathways in Exosomes Biogenesis, Secretion and Fate 2013).
2. La formation des ILV et la sélection du cargo moléculaire de l'exosome peuvent également être réglementées par un mécanisme indépendant d'ESCRT impliquant des microdomaines membranaires enrichis en tétraspanines ( formation des ILV par la voie dépendante des tétraspanines).
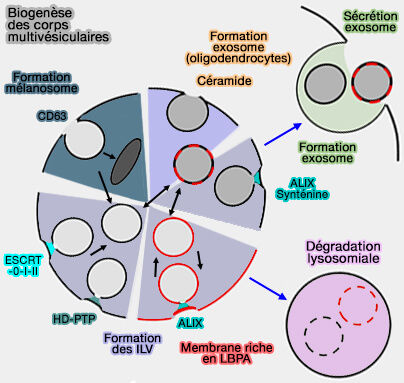
(Figure : vetopsy.fr d'après Gruenberg)
a. Les changements dans le niveau d'expression de CD9, CD81 et CD63 affectent la quantité totale d'exosomes sécrétés (Nanoparticle analysis sheds budding insights into genetic drivers of extracellular vesicle biogenesis 2016).
b. L'expression des tétrastanines TSPAN8 et CD63 affecte également la composition moléculaire des exosomes (Current knowledge on exosome biogenesis and release 2018).
Les tétraspanines interviennent également dans la formation des LRO (The tetraspanin CD63 regulates ESCRT-independent and dependent endosomal sorting during melanogenesis 2011).
c. Les acides nucléiques (ARNm, mi-ARN, ADN et ARN non codants) sont triés en exosomes à l'aide de protéines de liaison à l'ARN.
Le complexe ESCRT-II, ainsi que le microdomaine médié par la tétraspanine, le complexe de silençage induit par les mi-ARN (miRISC) et l'argonaute phosphorylé 2 (AGO2), reconnaissent des motifs d'ARN spécifiques et facilitent l'encapsulation de ces derniers par des exosomes (Shedding light on the cell biology of extracellular vesicles 2018).
3. Une voie dépendante d'ALIX et du complexe ESCRT favorise le tri et la livraison des tétraspanines aux exosomes (ALIX- and ESCRT-III–dependent sorting of tetraspanins to exosomes 2020).
- Une forme active de la protéine ALIX associée à l'ESCRT recrute efficacement les protéines ESCRT-III dans les endosomes.
- Ce recrutement, indépendamment des autres ESCRT, nécessite de l'acide lysobisphosphatidique (LBPA ou BMP).
Remarque : les exosomes sécrétés par les oligodendrocytes peuvent se former de manière dépendante des céramides, mais indépendante de l'ALIX et de l'ESCRT (Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes 2008).
Rôle des lipides membranaires des MVB
La composition membranaire des MVE/MVB, lors de la formation de l'ILV et de la fusion des MVE/MVB avec la membrane plasmique, souligne le rôle important joué par des lipides dans la biogenèse des exosomes (Extracellular vesicles: Lipids as key components of their biogenesis and functions 2018).

Les lipides spécifiques impliqués dans la formation des ILV favorisent la formation de microdomaines lipidiques et l'invagination membranaire (Lipid Rafts in Exosome Biogenesis 2020).
Sphingolipides : céramides et sphingosine-1-phosphate
1. Les céramides, qui font partie des sphingolipides, sont essentiels pour la courbure de la membrane et le bourgeonnement des vésicules (Role of sphingolipids in the biogenesis and biological activity of extracellular vesicles 2018).
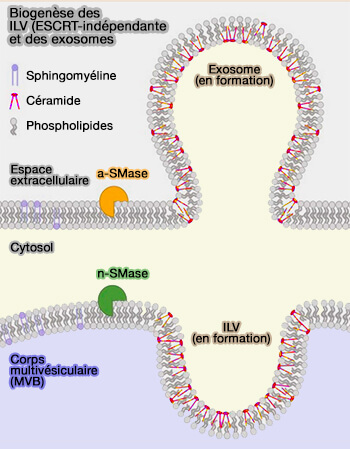
(Figure : vetopsy.fr d'après Verderio et coll)
Les sphingomyéline phosphodiestérases sont des enzymes qu'on appelle aussi sphingomyélinases (SMases).
- Les sphingomyélines représentent 85 % des sphingolipides et sont la source principale de céramide et de phosphocholine.
- Elles ont la propriété d'agir selon le pH.
Les céramides et les sphingomyélines sont traités dans des chapitres spéciaux
2. L'activation des récepteurs de la sphingosine 1-phosphate (S1PR) couplés aux Gi, protéines G inhibitrices, une des familles principales associées à un récepteur couplé aux protéines G (GPCR), sur les MVE/MVB, en particulier S1PR1, est essentielle pour le tri des cargos dans les ILV destinées à la libération d'exosomes (Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes 2013).
La sphingosine 1-phosphate est produite par la phosphorylation de la sphingosine catalysée par deux isoformes de sphingosine kinases (SK1 et SK2, EC 2.7.1.91), et l'activation de ses récepteurs S1P2R peut promouvoir la tumorigenèse (Sphingosine 1-phosphate and Cancer 2018).
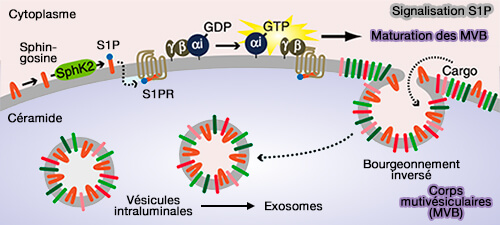
(Figure : vetopsy.fr d'après Kajimoto et coll)
3. PLP (ProteoLipid Protein ou lipophiline), dans les oligodendrocytes, est transférée dans des sous-domaines distincts de la membrane endosomale, puis les domaines associés aux exosomes sont transférés dans la lumière de manière indépendante du complexe ESCRT, ce qui nécessite également le céramide (Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes 2008).
Remarque : le mécanisme médié par les céramides peut être relativement indépendant de la biogenèse des ILV dépendantes de l'ESCRT.
Toutefois, le tri des cargos médié par l'ESCRT peut entrer en synergie avec la courbure lipidique induite par les céramides pour produire des vésicules qui partagent à la fois des mécanismes dépendants et indépendants de l'ESCRT (Extracellular vesicles: unique intercellular delivery vehicles 2017).
Glycérophospholipides
1. L'acide phosphatidique (PA) est caractérisé par une petite tête polaire négative qui génère une courbure membranaire négative (PLD2 is enriched on exosomes and its activity is correlated to the release of exosomes 2004).

Vous pouvez lire : Implication de l’acide phosphatidique dans le trafic membranaire : rôle et régulation de la phospholipase D au cours de la phagocytose et de l’exocytose régulée (2019).
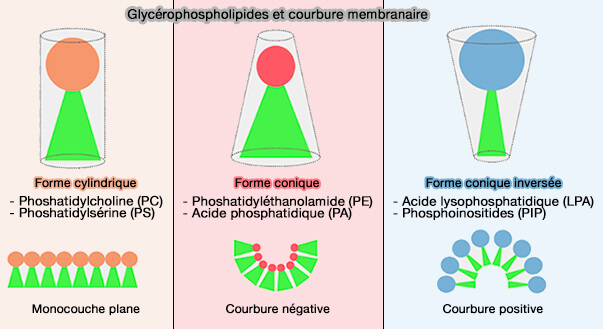
(Figure : vetopsy.fr d'après Tanguy)
Les enzymes productrices de PA régulent la libération d'exosomes dans plusieurs lignées cellulaires, i.e. :
- la phospholipase D (Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2 2014),
- la diacylglycérol kinase (Diacylglycerol kinase α regulates the formation and polarisation of mature multivesicular bodies involved in the secretion of Fas ligand-containing exosomes in T lymphocytes 2011).
2. Les lysophospholipides, i.e. phospholipides dont l'un des résidus d'acide gras estérifiant un hydroxyle du glycérol a été hydrolysé par une phospholipase, peuvent aussi courber les membranes.
Trafic et libération des exosomes
Vue d'ensemble
Les endosomes tardifs/endosomes ou corps multivésiculaires (MVE/MVB) ont un rôle central dans les mécanismes cellulaires et moléculaires régulant la libération, l'absorption et la fonction des exosomes.
1. Les endosomes tardifs peuvent fusionner ( exocytose lysosomale) :
- soit avec les lysosomes,
- soit avec les membranes plasmiques.
2. Les MVE/MVB fusionnent avec la membrane plasmique pour libérer leurs exosomes (Shedding light on the cell biology of extracellular vesicles 2018 et Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes 2016).
3. Les lysosomes peuvent aussi fusionner avec les membranes plasmiques ( exocytose lysosomale et membrane plasmique).
Bien que la fusion avec les lysosomes conduise à la destruction du contenu des endosomes, il semble que les exosomes, présents dans les lysosomes, sont protégés contre la dégradation et sont libérés par exocytose lysosomale.
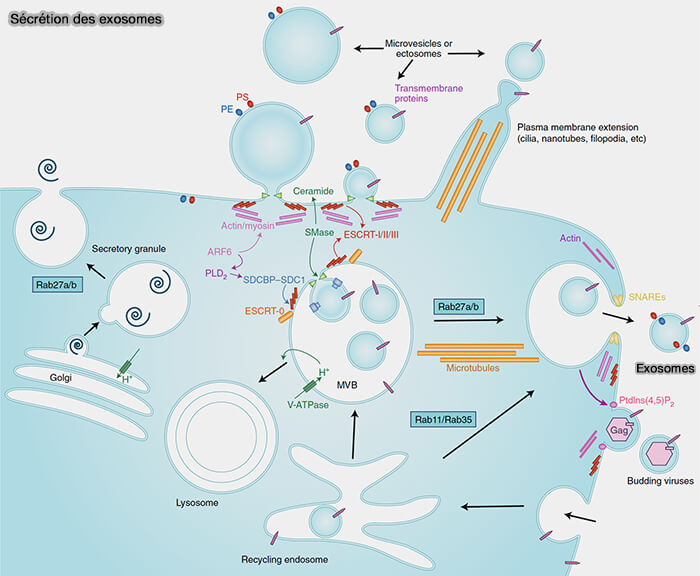
(Figure : vetopsy.fr d'après Mathieu et coll)
Molécules impliquées
1. Les petites GTPases Rab sont impliquées dans le trafic des MVE/MVB vers la membrane plasmique et leur amarrage pour la libération d’exosomes.
- a. Ce serait le cas de Rab27 (Rab27a and Rab27b control different steps of the exosome secretion pathway 2010).
Rab27a/b est aussi impliquée dans la sécrétion de corps de Weibel-Palade et de granules sécrétoires dérivés de l'appareil de Golgi ( transport par le cytosquelette d'actine dans le cortex cellulaire).
b. D'autres Rab seraient impliquées.
- Rab7 pour la formation des endosomes tardifs,
- Rab11 et Rab35, qui participent aussi au recyclage des cargos endocytosées non dépendantes de la clathrine ( endosomes de recyclage).
2. Les céramides générés par les SMases et le complexe ESCRT favorisent la formation de vésicules à la fois au niveau de la membrane plasmique et à l’intérieur des MVE/MVB.
- Arf6 et la dépolymérisation du cytosquelette d’actine sont nécessaires pour la sécrétion de vésicules extracellulaires dans les deux localisations.
- L’externalisation de la phosphatidyléthanolamine (PE) et de la phosphatidylsérine (PS) peut être plus spécifiquement impliquée dans la sécrétion des EV dérivée de la membrane plasmique et dans l’acidification médiée par la V-ATPase des MVE/MVB.
3. Les protéines SNARE sont impliquées dans les fusions et il n'est pas facile de savoir celles qui sont impliquées dans la fusion des MVE/MVB avec la membrane plasmique.
Autres vésicules extracellulaires : ectosomes
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation