


Autophagie
Protéines ATG (AuTophaGy-related)
ATG4
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Lors de l'autophagie, ATG4 est nécessaire pour lipider et délipider ATG8/LC3/GABARAP
ATG4 est nécessaire à ATG8 pour :
- initier la lipidation, en clivant pro-ATG8/LC3 pour exposer sa glycine N-terminale et former ATG8/LC3-I (
 système ATG8),
système ATG8), - la délipidation en clivant le lien entre Atg8 et la phosphatidyléthanolamine (PE) pour la fusion de l'autophagosome avec le lysosome (Atg4 in autophagosome biogenesis 2017).
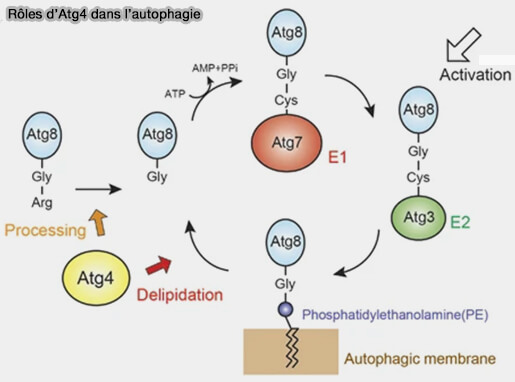
(Figure : vetopsy.fr d'après Maruyama et Noda)
Structure d'Atg4
1. Atg4 est conservé au cours de l’évolution de la levure aux mammifères.
- La levure n’a qu’un seul gène Atg4.
- Les mammifères possèdent quatre ATG4, i.e. Atg4A/autophagine-2, Atg4B/autophagine-1, Atg4C/autophagine-3 et Atg4D/autophagine-4 (Human Autophagins, a Family of Cysteine Proteinases Potentially Implicated in Cell Degradation by Autophagy 2003).
L’activité de la protéase est nettement différente entre les homologues Atg4 et la sensibilité de chaque homologue Atg4 est également différente parmi les homologues Atg8 (Kinetics Comparisons of Mammalian Atg4 Homologues Indicate Selective Preferences toward Diverse Atg8 Substrates 2010).

Vous pouvez lire : Autophagy-regulating protease Atg4: structure, function, regulation and inhibition (2018).
2. La structure d'Atg4 contient deux domaines :
- le domaine catalytique conservé parmi les protéases à cystéine de la famille des papaïnes,
- le domaine des doigts courts (short fingers domain) inséré dans le domaine catalytique et est unique à Atg4.
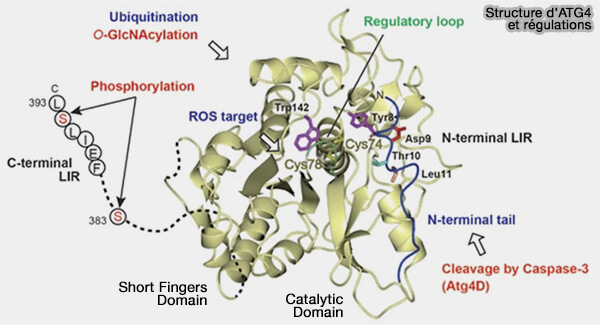
(Figure : vetopsy.fr d'après Maruyama et Noda)
Liaison Atg4/Atg8
Atg4 reconnaît Atg8 à l’aide d’au moins deux mécanismes :
- l’un utilise le motif LIR situé au niveau des queues flexibles N- et C-terminales,
- l’autre utilise le corps enzymatique (The structure of Atg4B–LC3 complex reveals the mechanism of LC3 processing and delipidation during autophagy 2009).
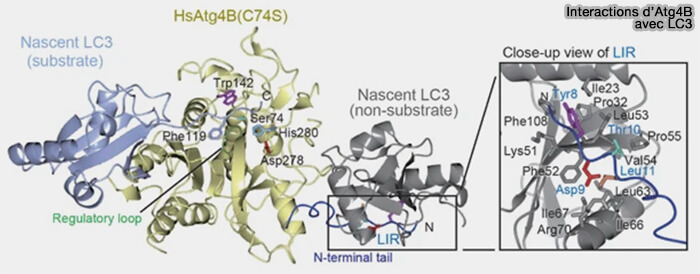
(Figure : vetopsy.fr d'après Maruyama et Noda)
Délipidation d'Atg8 par Atg4
1. Les membranes autophagiques LC3 positives formées pendant la déprivation persistent pendant environ 10 à 20 min après que la LC3 a commencé à s'associer à ces structures (Mechanisms and Pathophysiological Roles of the ATG8 Conjugation Machinery 2019).
a. Pendant cette période, le LC3/GABARAP lié à la membrane externe est important pour la maturation des autophagosomes en se liant à différentes protéines contenant des domaines LIR ( déplacement et attache des autophagosomes) :
- les domaines FYVE et FYCO1 qui facilitent l'interaction des autophagosomes avec les moteurs des microtubules orientés vers leur extrémité (+),
- le domaine PH de PLEKHM1 impliqué dans la fusion des autophagosomes aux lysosomes (PLEKHM1 Regulates Autophagosome-Lysosome Fusion through HOPS Complex and LC3/GABARAP Proteins 2015).
b. La délipidation LC3/GABARAP doit donc être strictement réglementée dans le temps et/ou l'espace pour empêcher leur élimination prématurée.
ATG4 rompt la liaison entre la glycine et la phosphatidyléthanolamine (PE).
Remarque : une libération très rapide de LC3 est effectuée par RavZ, une protéase anti-autophagie exprimée par des bactéries comme Legionella pneumophila qui entraîne la libération accélérée et irréversible de LC3 et de GABARAP à partir des membranes autophagiques (The Legionella Effector RavZ Inhibits Host Autophagy Through Irreversible Atg8 Deconjugation 2012).
Par contre, RavZ clive LC3-PE au-dessus de la glycine.
2. Un taux élevé d'espèces réactives de l'oxygène (ROS) serait nécessaire pour faciliter l'assemblage des autophagosomes en supprimant la délipidation de l'ATG8.
- En particulier, H2O2 s'accumule dans les cellules pendant les déprivations (Reactive oxygen species are essential for autophagy and specifically regulate the activity of Atg4 2007).
- Or, ATG4A et ATG4B contiennent des résidus de cystéine sujets à l'oxydation dans leurs sites actifs.
- Le stress oxydatif peut également provoquer une oxydation directe des SH, i.e. thiols d'ATG3 et d'ATG7, et inhibe leur activité dans la lipidation de la LC3 (Oxidation of Atg3 and Atg7 mediates inhibition of autophagy 2018).
3. Cependant, l'autophagie basale semble se produire de manière plus ou moins constitutive dans les cellules de mammifères, et la formation d'autophagosomes n'est pas limitée aux périodes de stress prolongé.
a. Tous les membres de la famille ATG4 contiennnent un motif LIR C-terminal.
b. En outre, l'ATG4B comprend un LIR N-terminal conservé qui a deux rôles (ATG4B contains a C-terminal LIR motif important for binding and efficient cleavage of mammalian orthologs of yeast Atg8 2017) :
- stabiliser le pool de protéines de la famille GABARAP solubles en empêchant leur dégradation par une voie de renouvellement indépendante de l'autophagie,
- soutenir la délipidation, par exemple de GABARAPL1-PE, réduit d'un facteur 4, et LC3B-PE complètement supprimé (Delipidation of mammalian Atg8-family proteins by each of the four ATG4 proteases 2018).
c. Cependant, la mutation du LIR C-terminal et N-terminal n'a aucun effet sur l'amorçage, celle du LIR N-terminal a seulement un effet modeste sur la délipidation (Delipidation of mammalian Atg8-family proteins by each of the four ATG4 proteases 2018).
4. De plus, la protéase ATG4 et le substrat d'ATG8 sont soumis à des modifications post-traductionnelles qui permettent un contrôle temporel strict sur l'élimination des protéines de la famille ATG8.
Modifications post-traductionnelles d'ATG4
Vous pouvez lire : Modifications post-Translational Modifications of ATG4B in the Regulation of Autophagy (2022).
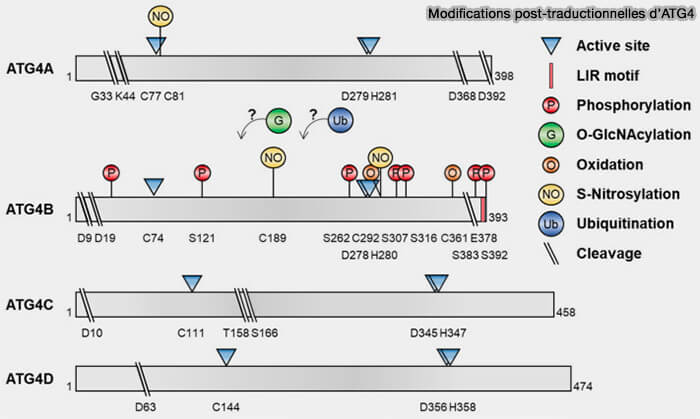
(Figure : vetopsy.fr d'après Park et coll)
Outre la phosphorylation, l'Atg4 est contrôlée par oxydation, ubiquitination, O-GlcNacylation et S-nitrosylation, permettant potentiellement des contrôles variables en fonction de l’état de stress cellulaire ( tableau).
1. La phosphorylation d’ATG4B sur S383 et S392 augmente son activité hydrolase envers LC3 conjuguée à la membrane et est importante pour un flux autophagique approprié, tandis que l’amorçage de LC3B ne semblait pas affecté par la phosphorylation d’ATG4B dans les cellules (ATG4B (Autophagin-1) Phosphorylation Modulates Autophagy 2015).
2. Le complexe ULK/Atg1 peut inhiber l'activité d'Atg4/ATG4B en phosphorylant les sérines d'Atg4, S307 pour la levure et S316 pour les mammifères (Atg4 proteolytic activity can be inhibited by Atg1 phosphorylation 2017).
a. Ainsi, l'accumulation locale d'Atg1/ULK1 à des phagophores en expansion va naturellement supprimer les protéines ATG4 actives.
- Le pool Atg8-PE est essentiel pour la formation d'autophagosomes et est partiellement protégée par la phosphorylation d'Atg4 par Atg1.
- Cependant, Atg8 nouvellement synthétisé est façonné constitutivement par Atg4.
b. La protéine phosphatase 2A (PP2A)-PP2R3B peut éliminer cette phosphorylation inhibitrice (A reversible phospho-switch mediated by ULK1 regulates the activity of autophagy protease ATG4B 2017).

Un commutateur kinase/phosphatase étroitement régulé semble contrôler l'activité de l'ATG4B et probablement d'autres protéines ATG4.
3. D'autres kinases peuvent phosphoryler ATG4 (ATG4B (Autophagin-1) Phosphorylation Modulates Autophagy 2015).
Remarque : la phosphorylation par TBK1 de LC3C et GABARAPL2 régule leur délipidation en perturbant spécifiquement la capacité des protéines ATG4 à reconnaître ces substrats (TBK1‐mediated phosphorylation of LC3C and GABARAP‐L2 controls autophagosome shedding by ATG4 protease 2020).
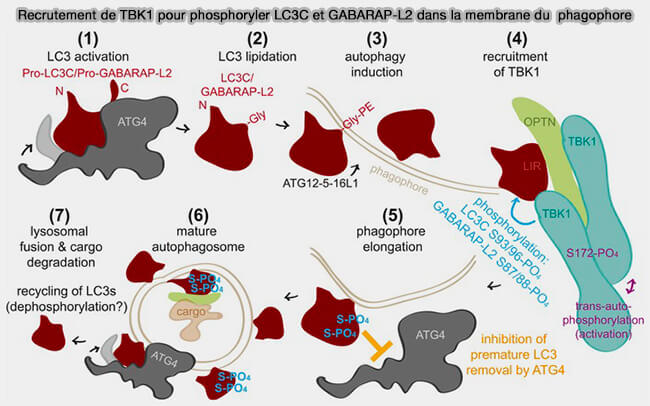
(Figure : vetopsy.fr d'après Herhaus et coll)
Retour à Atg8
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation