


Autophagie
Macroautophagie
Maturation des autophagosomes
Régulation
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La maturation des autophagosomes est régulée au niveau des SNARE, au niveau des phosphoinositides et des attaches et au niveau des gènes.
Ce chapitre est entièrement tiré de : Machinery, regulation and pathophysiological implications of autophagosome maturation (2021).
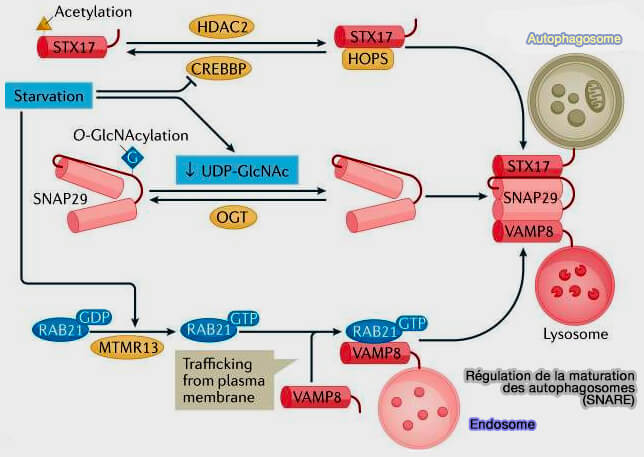
Régulation de la maturation au niveau des SNARE
1. Le domaine SNARE de la syntaxine 17 (STX17) est modifié par acétylation, un processus contrôlé par l'acétyltransférase CREBBP/CREBBP/CBP (CREB-Binding) et par la désacétylase HDAC2 (Acetylation of STX17 (syntaxin 17) controls autophagosome maturation 2021).
- La dénutrition inactive CREBBP et STX17 est alors désacétylée.
- STX17 interagit plus fortement avec le complexe HOPS et SNAP-29 et favorise ainsi la fusion autophagosome/lysosome.
(Figure : vetopsy.fr d'après Zhao et coll)
2. SNAP-29 est O-GlcNAcylé par l'OGT (O-GlcNAcylated by O-linked β-N-acetylglucosamine (O-GlcNAc) transferase).
- Cette modification diminue l'assemblage de SNAP29 avec les complexes SNARE.
- La dénutrition diminue la concentration intracellulaire de l'UDP-GlcNAc et donc la O-GlcNAcylation, ce qui facilite l'assemblée du complexe trans-SNARE pour la maturation de l'autophagosome (O-GlcNAc-modification of SNAP-29 regulates autophagosome maturation 2014).
3. MTMR13 (Myotubularin-related protein 13), la GEF (Guanine nucleotide exchange factor) pour RAB21 endosomal, contrôle le trafic de VAMP8 de la membrane vers les endosomes tardifs/lysosomes (Starvation-induced MTMR13 and RAB21 activity regulates VAMP8 to promote autophagosome–lysosome fusion 2015).
Lors de dénutrition, MTMR13 active RAB21, ce qui favorise par la suite la translocation de VAMP8 pour promouvoir la fusion des endosomes/lysosomes avec des vacuoles autophagiques.
Régulation de la maturation au niveau
des phosphoinositides et des attaches
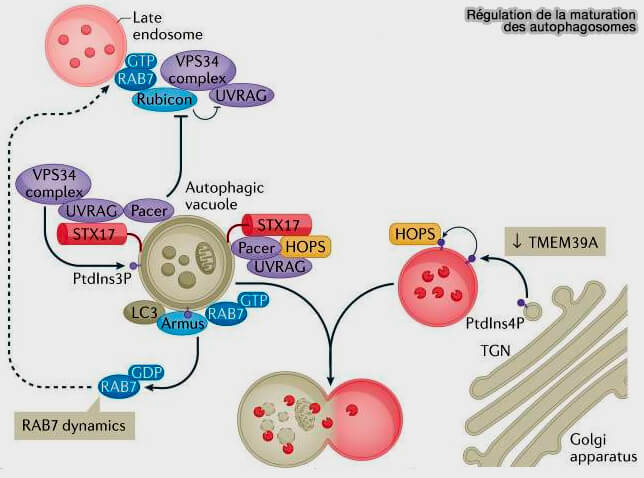
1 PI(3)P sur des vacuoles autophagiques, généré par UVRAG, favorise l’activation du complexe de nucléation PI3KC3-CI contenant VPS34, facilite la maturation des autophagosomes par le recrutement du complexe HOPS.
a. Rubicon interagit avec le complexe UVRAG-VPS34 et régule négativement sa fonction.
b. Pacer est ciblée sur les vacuoles autophagiques STX17 et PI(3)P, antagonise Rubicon et recrute le UVRAG-VPS34 (The Autophagy Protein Pacer Positively Regulates the Therapeutic Potential of Mesenchymal Stem Cells in a Mouse Model of DSS-Induced Colitis 2022).
Pacer et UVRAG peuvent également recruter le complexe HOPS.
(Figure : vetopsy.fr d'après Zhao et coll)
2. Armus, la GAP (GTPase-Activating Protein de RAB7, interagit avec LC3 et PI(3)P, favorise le recyclage de RAB7 des membranes autophagiques aux endosomes pour faciliter leur maturation en endosomes tradifs (LE)/lysosomes.
(Figure : vetopsy.fr d'après Zhao et coll)
3 La déplétion de la TMEM39A endoplasmique augmente la concentration de PI(4)P sur les endosomes tardifs/lysosomes, via l'inhibition du trafic Golgi ➞ RE de SAC1, la PI(4)P phosphatase, ce qui favorise le recrutement du complexe HOPS et améliore la fusion autophagosome/lysosome.
Remarque : GABARAP pourrait recruter une PI4-kinase, i.e. PI4KIIA aux autophagosomes, conduisant à la génération de PI(4)P qui favorise la fusion des autophagosomes avec le lysosome.
Régulation de la
maturation au niveau
des gènes
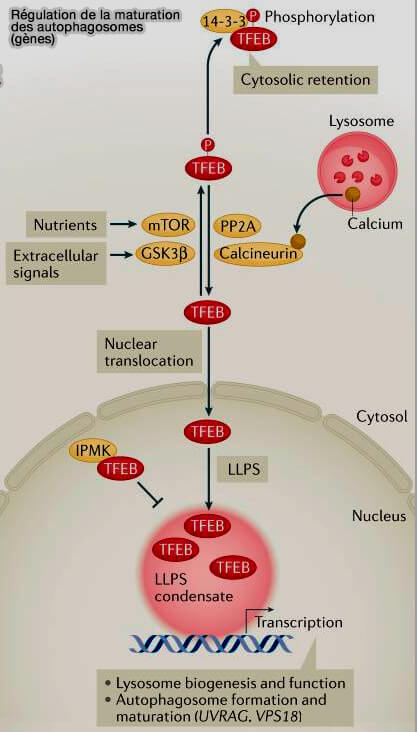
1. Le facteur de transcription TFEB et son homologue TFE3 active l'expression de gènes impliqués dans l'autophagie, i.e. gènes impliqués dans le trafic, dans la synthèse d'UVRAG, de VPS38 et dans la biogenèse des lysosomes ( régulation transcriptionnelle de la biogenèse des lysosomes).
régulation transcriptionnelle de la biogenèse des lysosomes).
1. Le transport nucléaire de TFEB est réglementé par ses niveaux de phosphorylation.
- Diverses kinases, telles que mTORC1, en aval des nutriments, et GSK-3β (Glycogène Synthase Kinase 3), en aval de PKC (Protéine Kinase C) de signalisation en réponse à divers signaux extracellulaires, phosphorylent TFEB pour empêcher son importation vers le noyau.
- La protéine phosphatase 2A (PP2A) et la calcineurine, qui est activé par la libération de calcium à partir des lysosomes, déphosphoryle TFEB pour faciliter sa translocation vers le noyau.
2. L'activité de TFEB est également contrôlée par la séparation de phase liquide-liquide (LLPS), en vertu de laquelle TFEB forme des condensats pour promouvoir la transcription des gènes (Liquid–liquid phase separation in autophagy 2020).
La protéine nucléaire de l'inositol polyphosphate multikinase (IPMK) se lie directement à TFEB et inhibe la formation des condensats de TFEB.
Fusion des autophagosomes
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation