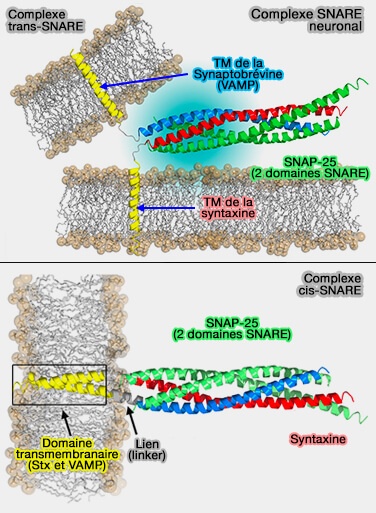
L'assemblage du complexe trans-SNARE nécessite plusieurs protéines SNARE, i.e. trois ou quatre, nécessaires à la formation du faisceau de 4 hélices α issues des domaines SNARE pour former un motif coiled-coil.
Le processus d'assemblage est un continuum qui produit :
un complexe trans-SNARE, i.e. les SNARE sur des membranes opposées,
Complexe SNARE neuronal
(Figure : vetopsy.fr d'après Fang et Lindau)
un complexe cis-SNARE, i.e. les SNARE sur la même membrane.
Vue d'ensemble de la formation du complexe trans-SNARE
Le pliage et l'assemblage directionnels et pas à pas des faisceaux à quatre hélices SNARE sont généralement appelés fermetures à glissière (ou fermetures éclair) SNARE (SNARE zippering).
2. En raison de la rigidité du lien en liaison avec les domaines SNARE et les TMD, la fermeture à glissière rapproche les TMD et les aligne finalement l'un avec l'autre lors de la fusion.
Les TMD exercent donc une force sur ces membranes.
Les TMD ont des fonctions mécaniques qui vont au-delà de l'ancrage passif du complexe SNARE générateur de force à la fusion des membranes.
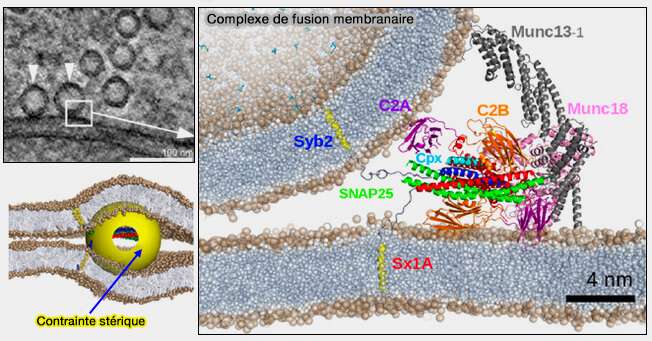
Complexe de fusion membranaire
(Figure : vetopsy.fr d'après Grubmüller et coll)
Cependant, on ne sait pas comment cette force est transmise aux extrémités C-terminales des TMD, i.e. partie qui doit agir activement sur le feuillet interne de la bicouche pour pouvoir par la suite produire un stade intermédiaire d'hémifusion, puis pour ouvrir et élargir un pore de fusion.
En effet, les SNARE avec un TMD tronqué ou avec une ancre de membrane lipidique, qui ne couvre que la moitié de la bicouche, sont inefficaces pour former des pores de fusion.
La force que les complexes SNARE peuvent exercer sur les intermédiaires de fusion dépend de la structure secondaire adoptée du lien de liaison reliant le domaine SNARE au TMD
.
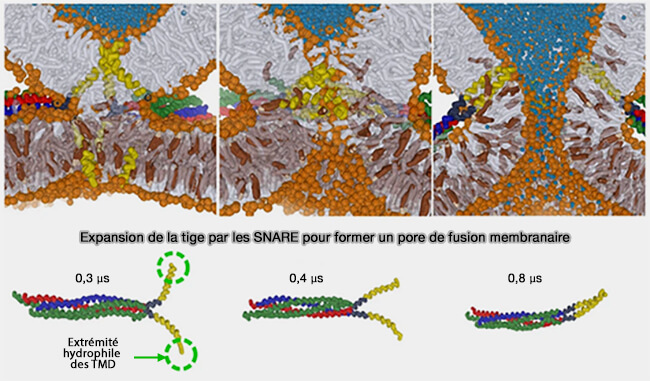
Expansion de la tige par les SNARE pour former un pore de fusion membranaire
(Figure : vetopsy.fr d'après Risselada et coll)
Étapes de la fermeture à glissière des domaines SNARE
1. La syntaxine 1 (Qa) et SNAP-25 (Qb/c avec ces deux domaines SNARE) forment un complexe t-SNARE binaire 1:1 (ou complexe Qabc), faisceau parallèle à trois hélices avec des terminaisons C désordonnées, i.e. couches +5 à + 8.
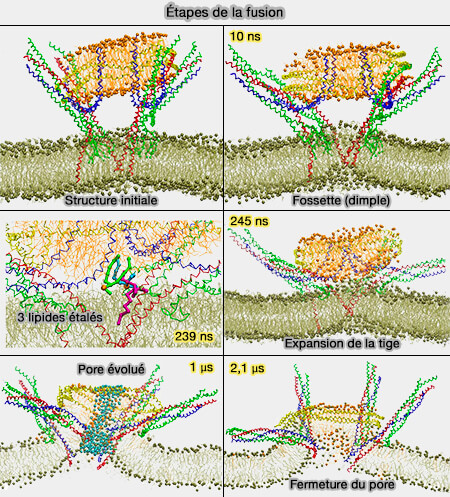
Étapes de la fusion
(Figure : vetopsy.fr d'après Sharma et coll)
Cependant, lorsque les trois SNARE sont mélangés simultanément, l'assemblage du complexe ternaire est beaucoup plus lent, i.e. nécessitant des dizaines de minutes à des heures.
c. Enfin, survient la reconfiguration du lien (linker) juxtamembranaire (LD).
Remarque : dans l'ensemble, l'assemblage d'un seul complexe synaptique SNARE produit une énergie totale d'environ 85 kBT, dont 68 kBT provenant de la fermeture éclair entre les t- et v-SNARE et 17 kBT provenant du repliement du complexe t-SNARE (Chaperoning SNARE Folding and Assembly 2021).
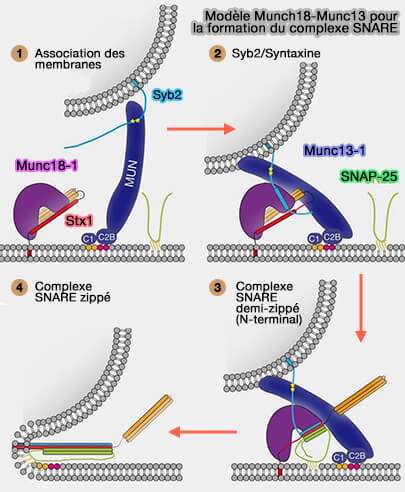
Modèle Munch18-Munc13
(Figure : vetopsy.fr d'après Wang et coll)
La fermeture éclair NTD est responsable de l'amarrage (docking) des vésicules synaptiques et de l'amorçage (priming) des SNARE pour la fusion déclenchée par Ca++.
La fermeture éclair CTD est étroitement couplée à la fusion de la membrane et à la libération des neurotransmetteurs.
Cette conformation arrête ainsi la progression de la fermeture à glissière SNARE et altère la transduction de force efficace des domaines SNARE vers les TMD.
Ce phénomène pourrait expliquer pourquoi des mutations dans la région du TMD de la synaptobrévine 2/VAMP2, qui améliorent en fait l'hélicité et la rigidité du lien dans les simulations moléculaires, réduisent néanmoins l'activité de fusion observée expérimentalement ( flexibilié conformationnelle des TDM des v-SNARE).
Une transduction de force efficace des complexes SNARE aux intermédiaires de fusion nécessite une formation régulée de la structure α-hélicoïdale, ce qui est le cas par le contrôle des protéines associées aux SNARE, i.e. les protéines SM (Sec1/Munc18) et les Protéines d'attache (tethering protein).
Le rapprochement des deux membranes nécessite un apport d'énergie important pour surmonter les forces électrostatiques répulsives.
1. Une des hypothèses les plus plausibles est que les protéines SNARE, par la formation du complexe trans-SNARE, libèrent de l'énergie pour courber la membrane ( courbure membranaire).
Étapes de la fermeture à glissière des complexes SNARE
(Figure : vetopsy.fr d'après Zhang)
Au fur et à mesure que les domaines SNARE s'enroulent, ils forment un faisceau à quatre hélices beaucoup plus serré et plus stable en forme de fermeture éclair.
Lors de la formation de la fermeture éclair, une fraction de l'énergie libérée par la liaison des domaines SNARE est stockée sous forme de contrainte de flexion moléculaire dans les motifs SNARE individuels.
Cette contrainte mécanique est supposée être stockée dans les liens (linker), régions de liaison semi-rigides entre les domaines transmembranaires (TMD) et le faisceau SNARE hélicoïdal.
2. La flexion énergétiquement défavorable est minimisée lorsque le complexe se déplace en périphérie où la courbure est plus importante ( complexe trans-SNARE et encombrement stérique).
Toutefois, les modèles moléculaires de la fusion conduite par les SNARE ne sont pas encore complètement connus !





 fusion et encombrement stérique).
fusion et encombrement stérique).