ApoER2 (apolipoprotein E Receptor 2 ou LRP8) et VLDLR (Very Low Density Lipoprotein Receptor) appartiennent à la famille des LDLR (Low-Density Lipoprotein Receptor)et joue un rôle majeur au niveau cérébral.
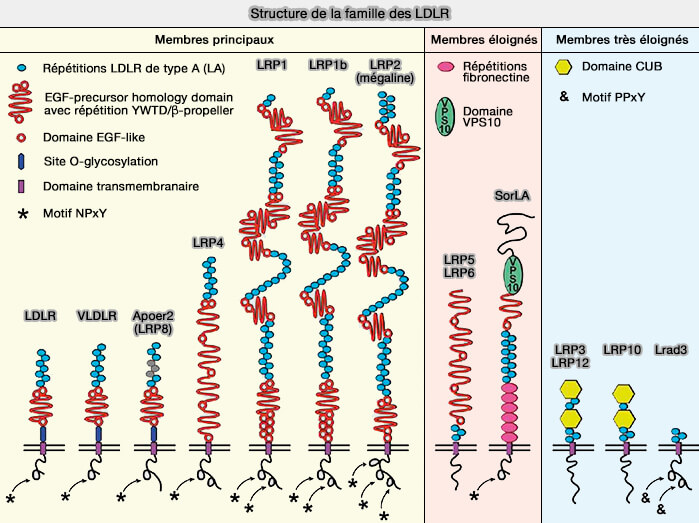
Structure de la famille des LDLR
(Figure : vetopsy.fr d'après Pohlkamp et coll)
Vue d'ensemble d'ApoER2 (LRP8) et VLDLR
ApoER2 (apolipoprotein E Receptor 2 ou LRP8) et VLDLR (Very Low Density Lipoprotein Receptor) sont principalement exprimés dans le cerveau.
1. Contrairement à LDLR, dont la structure est très proche, qui a une fonction très spécifique dans la régulation de l'homéostasie du cholestérol, ApoER2 et VLDLR participent à une multitude de voies et de modules fonctionnels différents.
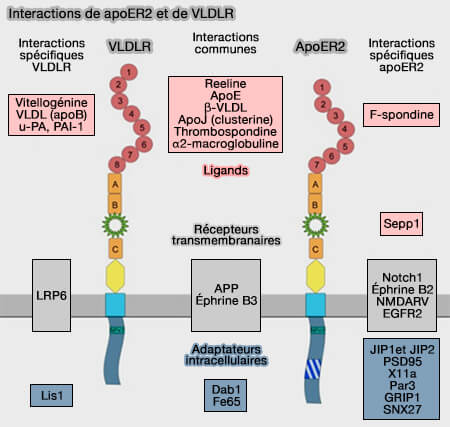
Les trois récepteurs se lient, entre autres, avec l'apoE.
avec de nombreux ligands, certains communs, d'autres non,
comme récepteurs de transduction du signal,
Interactions de ApoER2 et VLDLR
(Figure : vetopsy.fr d'après Dlugosz et Nimpf)
seuls ou ensemble comme dans la signalisation reeline, glycoprotéine neuromodulatrice de la matrice extracellulaire,
avec d'autres protéines adaptatrices membranaires ou non pour jouer le rôle de plates-formes de signalisation aux fonctions encore inconnues.
Leur structure générale est celle des membres de la famille LDLR, en particulier de LDLR, avec lequel ils partagent une liaision avec l'apoE. Nous ne soulignerons que leurs différences.
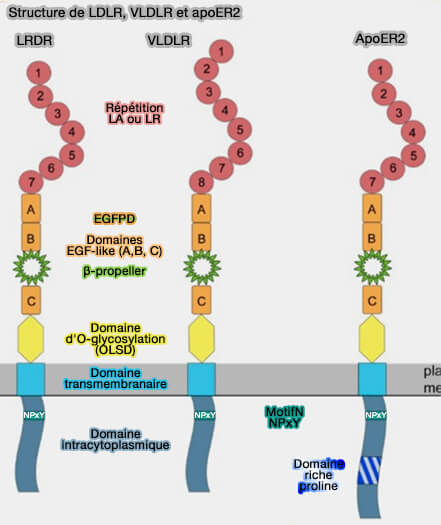
Structure de LDLR, VLDLR et ApoER2
(Figure : vetopsy.fr d'après Dlugosz et Nimpf)
ApoER2 en comprend sept dans sa longueur maximale ( épissage),
VLDLR huit.
2. ApoER2 contient un domaine cytoplasmique riche en proline (PRD) de 59 résidus, unique dans la famille des LDLR, qui se termine à 10 résidus de l'extrémité C-terminale.
Ce domaine, qui contient deux motifs de liaison SH3 potentiels, PxxP, ce qui suggère un rôle dans la transduction du signal et favorise les interactions de ApoER2 avec :
3. VLDLR et ApoER2 possède, comme LDLR, un domaine d'O-glycosylation (OLSD ou O-Linked Sugar Domain) de 47 résidus pour VLDLR, de 89 pour LRP8, localisé juste au-dessus du feuillet externe de la membrane plasmique
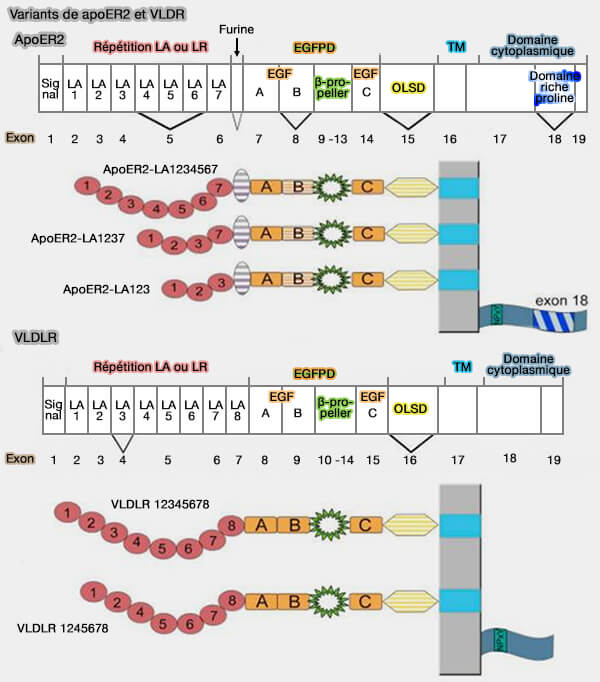
1. Dans le cerveau, humain, la forme prédominante est dépourvue de l'exon 5, codant pour les répétitions LA 4 à 6, i.e. le récepteur ne contient que les répétitions LR 1 à 3 et 7.
Variants de ApoER2 et VLDLR
(Figure : vetopsy.fr d'après Dlugosz et Nimpf)
2. Le domaine e d'O-glycosylation (OLSD ou O-Linked Sugar Domain) représente le site de moindre homologie entre ApoER2 et VLDLR.
Pour les deux récepteurs, les variants d'épissage contenant le domaine OLS sont fortement glycosylés et cette glycosylation inhibe le traitement protéolytique.
Seuls, LDLR, VLDLR et ApoER2 sont ciblés par IDOL.
Remarque : IDOL régulerait par son action sur l'action du VLDLR dans le système nerveux central pour le maintien de l'homéostasie énergétique du corps entier ( rôles de VLDR dans le métabolisme lipidique).
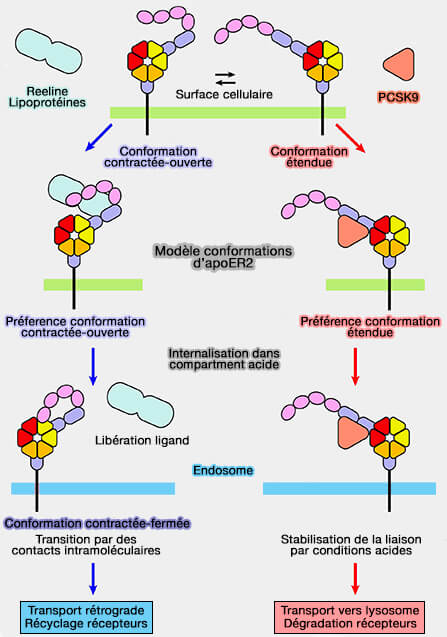
Conformations d'ApoER2
Figure : vetopsy.fr d'après Hirai et coll)
3. L'ApoER2 peut se présenter sous plusiuers formes, i.e. étendue, contractée ouverte et contractée fermée.
a. En complexe avec la reeline, il adopte la conformation contractée-ouverte. Puis lors de son internalisation, il adopte une conformation fermée-contractée pour libérer le ligand, comme le fait LDLR, et le récepteur est recyclé vers la surface cellulaire.




 endocytose clathrine-dépendante),
endocytose clathrine-dépendante),