


Lipides
Trafic non vésiculaire
Vue d'ensemble
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

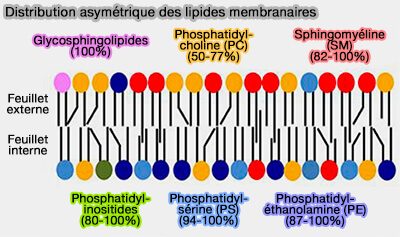
Les lipides sont distribués de manière très asymétrique dans différentes membranes cellulaires et une majorité de lipides utilise un transport non vésiculaire contrairement aux protéines.
Généralités sur trafic non vésiculaire des lipides
Vue d'ensemble et avantages
Les lipides sont distribués de manière très asymétrique dans différentes membranes cellulaires et une minorité de lipides réalisent leur distribution intracellulaire finale par sélection dans les membranes des vésicules de transport (Mechanisms of nonvesicular lipid transport 2021).
(Figure : vetopsy.fr d'après van Ijzendoorn et coll)
La composition lipidique de chaque membrane est précisément adaptée à ses fonctions, avec un système dédié de trafic lipidique intracellulaire pour réaliser différents mélanges lipidiques (Membrane lipids: where they are and how they behave 2008).
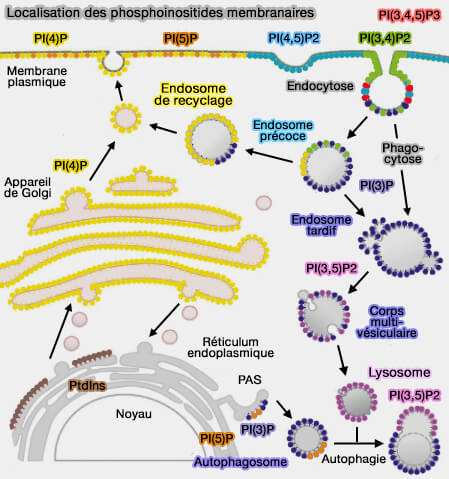
1. Le transport non vésiculaire peut fournir une grande quantité de lipides nécessaires à la biogenèse membranaire des organites qui ne peuvent pas les obtenir à partir du trafic vésiculaire.
- Les mitochondries (ou les chloroplastes), les goutelettes lipidiques (LD ou Lipid Droplet) ne disposent pas de la plupart des enzymes nécessaires à la fabrication de certains lipides nécessaires à leur biogenèse (Phospholipid ebb and flow makes mitochondria go 2020).
- Ces organites ne sont pas connectés au reste de la cellule par des voies de trafic vésiculaire et dépendent, donc de voies de trafic non vésiculaires pour obtenir ces lipides.
(Figure : vetopsy.fr d'après Cernikova et coll)
2. Le transport lipidique non vésiculaire permet de modifier, i.e. appauvrir ou enrichir, ou de maintenir le niveau approprié d'un lipide dans un organite ou un domaine d'un organite plus rapidement et plus précisément que ce qui est possible par le trafic vésiculaire ou dans des conditions de stress lorsque le trafic vésiculaire est compromis.
a. Il peut déplacer rapidement les lipides entre des compartiments spécifiques dans les cellules sans avoir à transférer également des protéines membranaires intégrales.
- Le trafic non vésiculaire du cholestérol fournit probablement une voie pour déplacer le cholestérol rapidement et efficacement vers les enzymes qui effectuent ces réactions sans perturber le trafic vésiculaire.
- Le transfert non vésiculaire des céramides du RE, où ils sont synthétisés, vers l'appareil de Golgi, où ils sont convertis en glycolipides et sphingolipides, peut réguler la production de ces lipides (
 biosynthèse des sphingophospholipides).
biosynthèse des sphingophospholipides).
(Figure : vetopsy.fr d'après Shrestra et coll)
b. Il permet la transmission d'un lipide dans le cadre d'une voie de signalisation ou de régulation.
Par exemple, le diglycéride (DAG) active la protéine kinase C (PKC) et les céramides servent de molécules de signalisation pour réguler la différenciation, la prolifération, la mort cellulaire programmée et l'apoptose.
3. Ce transfert intracellulaire diffère des systèmes de transfert de lipides à l'extérieur des cellules par les lipoprotéines, mais qui peuvent aussi utiliser certaines protéines de transfert de lipides (LTP).
Sites de contact
membranaire (MCS)
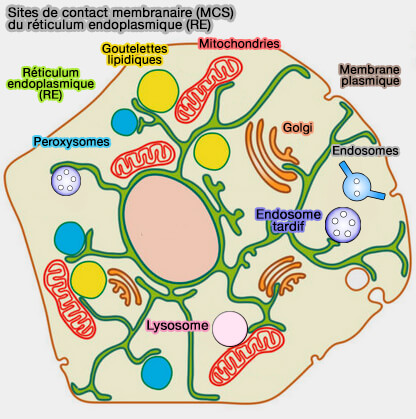
La plupart de ces transports se produisent probablement aux sites de contact membranaire (MCS) , où les organites sont étroitement apposés.
(Figure : vetopsy.fr d'après Inserm)
1. Les sites de contact membranaire (MCS) sont des régions d'apposition étroite (≤ 30 nm) entre deux organites, établissant des microdomaines qui permettent un échange de matériel.
Les progrès récents de la biologie des MCS ont révélé leurs rôles clés dans les échanges inter-organites.

Les sites de contact membranaire du réticulum endoplasmique (RE) sont étudiés dans des chapitres spécifiques.
- MCS avec les endosomes ou corps multivésiculaires (MVE/MVB) et les lysosomes,
- MCS RE/membrane plasmique,
- MCS RE/Golgi,
- MCS RE/mitochondries,
- MCS RE/peroxysomes,
- MCS RE/gouttelettes lipidiques.
2. Ces échanges ont été caractérisé pour le transport et la synthèse des lipides :
- le cholestérol,
- la phosphatidylcholine (PC) et la phosphoatidyléthanolamine (PE),
- les céramides.
Les MCS sont aussi impliqués dans la formation des gouttelettes lipidiques, i.e. Lipid Droplets ou LD (Lipid trafficking and membrane contact sites de l'inserm).
Modèles mécaniques de diffusion des lipides
Hypothétiquement, l'échange de lipides entre les membranes implique des transports passifs, i.e. ne nécessitant pas d'énergie (Lipid Trafficking sans Vesicles: Where, Why, How? 2010).
Remarque : les lipides peuvent être aussi transportés par un transport actif utilisant l'ATP comme dans les flippases, i.e. transporteurs ABC ou P4-ATPases, des lipoprotéines ou par trafic vésiculaire.
Diffusion simple
Les modèles d'échange passif de lipides, i.e. diffusion simple, impliquent un transfert spontané de lipides en l'absence d'un intermédiaire protéique, et ce selon le sens du gradient de concentration, i.e. elle est prédominante lors de la phase post-prandiale (Membrane Contact Sites: Complex Zones for Membrane Association and Lipid Exchange 2016).
1. L'échange par diffusion se produit lorsqu'un lipide est éjecté dans la phase aqueuse, suivie de sa diffusion et de sa réinsertion dans une membrane acceptrice.
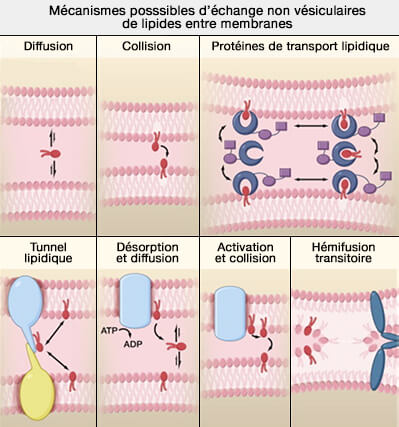
lipides entre membranes
(Figure : vetopsy.fr d'après Prinz)
2. L'échange lipidique par collision peut, en principe, se produire par l'adhésion de deux particules accompagnée d'un transfert de lipides à travers une couche polaire (partiellement) hydratée.
a. Cet échange direct de lipides lorsque les bicouches entrent en collision de manière transitoire, minimisant ainsi l'exposition au cytoplasme aqueux.
b. Certains auteurs incluent l'hémifusion qui impliquerait une collision plus persistante par lequel l'échange direct de lipides se produit par fusion entre les membranes du réticulum endoplasmique (RE) et la membrane plasmique (MP).
- L'intermédiaire d'hémifusion pourrait également être facilité par des protéines conduisant à une fusion incomplète, i.e. médiée par les protéines SNARE, par exemple, entre le R-SNARE Sec22p résidant dans le RE et un Q-SNARE localisé dans la MP.
- Cependant, on serait en présence de transport actif, nécessitant de l'énergie, même s'il n'y a pas de trafic vésiculaire.
Remarque : bien que l'on pense que l'hémifusion se produit de manière transitoire lors de la fusion membranaire, rien ne prouve que ce mécanisme soit utilisé pour déplacer les lipides entre les compartiments cellulaires qui ne fusionnent pas.
Diffusion facilitée
La diffusion facilitée, qui est aussi une diffusion passive, fait intervenir des protéines transmembranaires , i.e. des protéines de transfert de lipides (LTP) qui facilitent la vitesse de passage, i.e. elle est prédominante à de faibles concentrations (Mechanisms of nonvesicular lipid transport 2021).
Ces protéines de transfert de lipides (LTP) liées à la membrane ou solubles, qui ont la capacité de déplacer les lipides entre les membranes via des cavités hydrophobes qui protègent les lipides de l'environnement aqueux pendant le transport ( :
Les protéines de transfert de lipides (LTP) sont étudiées dans un chapitre spécifique.
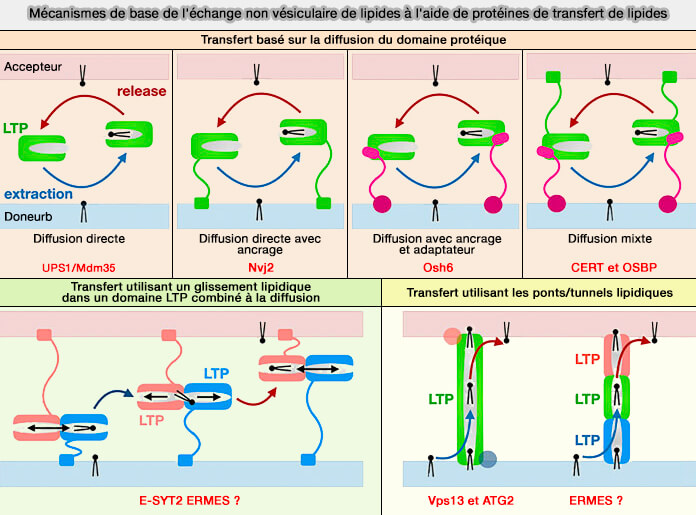
(Figure : vetopsy.fr d'après Egea)
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesGlycérophospholipidesSphingophospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation