


Transport des lipides
Lipoprotéines : récepteurs
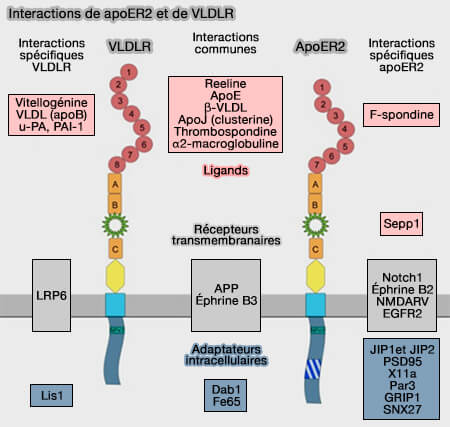
Récepteurs LDLR : ApoER2 (LRP8) et VLDLR
Fonctions
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Vue d'ensemble des lipides
- Acides gras
- Classification des lipides
- Cholestérol
- Glycérides
- Trafic non vésiculaire des lipides et LTP
- Digestion et absorption entérocytaire des lipides
- Lipoprotéines
- Gouttelettes lipidiques
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

ApoER2 (apolipoprotein E Receptor 2 ou LRP8) et VLDLR ((Very Low Density Lipoprotein Receptor) joue un rôle majeur au niveau cérébral avec ou sans la reeline.
Signalisation reeline
Signalisation reeline neuronale
ApoER2 et VLDLR sont principalement exprimés dans le cerveau où ils agissent comme des récepteurs non seulement pour l'apoE, mais aussi pour la reeline, une glycoprotéine neuromodulatrice de la matrice extracelluaire (ECM).
Son nom provient de la démarche titubante (reeling gait) des souris Reeler, homozygotes pour la mutation du gène RELN, codant cette protéine.
Vue d'ensenble

La reeline et ses rôles sont bien détaillés dans : The functions of Reelin in membrane trafficking and cytoskeletal dynamics: implications for neuronal migration, polarization and differentiation (2017).
1. La signalisation Reelin implique une voie centrale qui s'active lors de la liaison de la reeline à ses récepteurs, en particulier à ApoER2 et VLDLR.
- Cette liaison provoque la phosphorylation de la protéine adaptatrice Disabled-1 (Dab1) par Src/FYN.
- Dab1 phosphorylé (pDab1) est un hub dans la cascade de signalisation, à partir de laquelle plusieurs autres voies en aval divergent reflétant les différents rôles de la reeline.
Ces voies impliquent :
- la dynamique de l'actine et des microtubules,
- le trafic membranaire via la régulation de l'activité de les petites GTPases, i.e. Rho et Rap (Ras proximate 1, et les molécules impliquées dans la polarisation cellulaire,
- la modulation de la fusion spontanée des vésicules synaptiques (
 VAMP7 et fusion membranaire)
VAMP7 et fusion membranaire)

Les fonctions de la reeline sont complexes et le mode d'action de cette cascade de signalisation in vivo aux niveaux cellulaire et moléculaire est encore obscur.
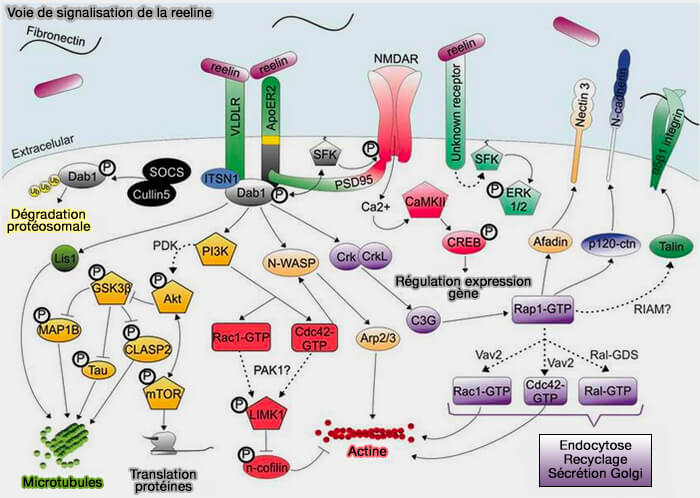
(Figure : vetopsy.fr d'après Dlugosz et Nimpf)
2. La signalisation initiée par la liaison de la reeline à ApoER2 et VLDLR joue un rôle essentiel dans le cerveau.
Pour tout savoir sur la reeline et le cerveau, lire : Reelin Functions, Mechanisms of Action and Signaling Pathways During Brain Development and Maturation (2020)
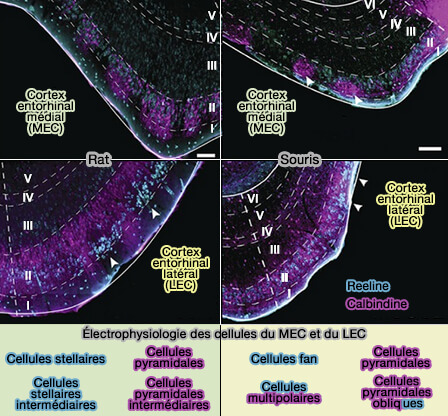
(Figure : vetopsy.fr d'après Witter et Mott)
a. Lors du développement précoce du cortex cérébral, la reeline régule la migration radiale des neurones corticaux (Reelin and brain developpment 2003 et Role for Reelin in stabilizing cortical architecture 2010).
- Elle est exprimée et sécrétée par les cellules Cajal-Retzius, appelées aussi cellules horizontales de Cajal, et module le cytosquelette et la mobilité des neurones migrants (The functions of Reelin in membrane trafficking and cytoskeletal dynamics: implications for neuronal migration, polarization and differentiation 2017).
- Elle assure une lamination corticale, hippocampique et cérébelleuse appropriée (Role of Reelin in the development and maintenance of cortical lamination 2009).
b. Lors du développement plus tardif, la reeline favorise la maturation des dendrites et des épines dendritiques (The Reelin Signaling Pathway Promotes Dendritic Spine Development in Hippocampal Neurons 2008).
c. Elle égule la migration continue des neuroblastes générés dans les sites de neurogenèse adultes comme les aires sous-ventriculaire (SVZ) de la paroi des ventricules cérébraux latéraux et sous-granulaire du gyrus denté.
d. Dans le cerveau mature, elle est impliquée dans la modulation de la plasticité synaptique en améliorant l'induction et le maintien de la potentialisation à long terme (Apoer2: A Reelin Receptor to Remember 2005).
Remarque : la reeline est retrouvée également dans un grand d'organe, i.e. foie, glande thyroïde, glande surrénale, trompes de Fallope, sein… ( The human protein atlas).
Structure et protéolyse
La structure de la reeline (3461 residus) est détaillée dans : Structural Insights into Reelin Function: Present and Future (2016).
1. La reeline comprend :
- un domaine signal,
- un domaine reeler, i.e.F-sp (F-spondin homology domain),
- un domaine H unique, séparé en trois régions, x, y et z,
- huit répétitions reeline, i.e. R1 à R8, composées chacune de deux sous-répétititions (A and B) entourant un motif EGF-like,
- un domaine CTR (C-terminally RR8) qui contient une séquence de résidus basiques.
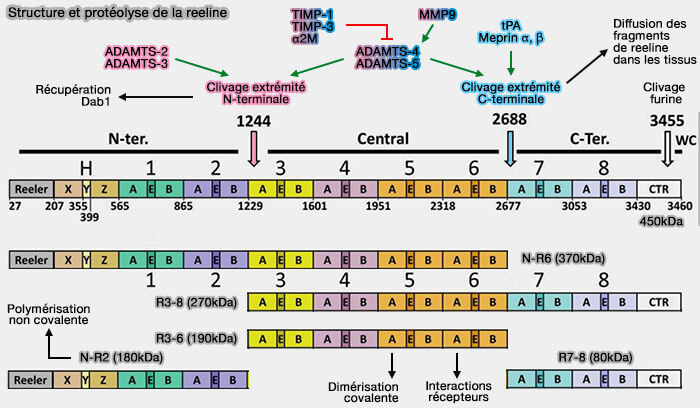
(Figure : vetopsy.fr)
2. La signalisation de la reeline est étroitement régulée par son clivage protéolytique en deux sites, ce qui donne naissance à cinq fragments différents, i.e. 370, 270, 190, 180 et 80 kDa, qui ont diverses fonctions (revue dans Reelin Functions, Mechanisms of Action and Signaling Pathways During Brain Development and Maturation 2020).
Un autre clivage possible, i.e. WCT par la furine [R-X-(R/K)-R] entre les résidus 3452 et 3455 (3452RRRR3455SLRRYP3461), pourrait libérer les six derniers acides aminés dont la fonction est inconnue.
3. La reeline est sécrétée sous forme de dimère et Cys2101 est crucial pour la formation de la liaison disulfure, i.e. le mutant reeline dépourvu de Cys2101 n'est pas capable d'induire une signalisation intracellulaire en aval, i.e. la phosphorylation de Dab1 (Functional Importance of Covalent Homodimer of Reelin Protein Linked via Its Central Region 2011).
4. ApoER2 se lie à R5-R6 de la reeline se lie avec une grande affinité à la première répétition LR (LA1), mais aussi à LA2 et EGFPD (Structural basis for ligand capture and release by the endocytic receptor ApoER2 2017).
a. La liaison est dépendante du calcium et peut être inhibée par l'apoE.
b. Les variants épissés d'ApoER2 se lient auxs fragments de reeline avec différentes affinités et sont exprimées de manière différentielle de manière spatio-temporelle dans le cerveau de souris en développement (Differential Action of Reelin on Oligomerization of ApoER2 and VLDL Receptor in HEK293 Cells Assessed by Time-Resolved Anisotropy and Fluorescence Lifetime Imaging Microscopy 2019).
- Le fragment ECD soluble d'ApoER2 agit comme un récepteur négatif dominant (A secreted soluble form of ApoE receptor 2 acts as a dominant-negative receptor and inhibits Reelin signaling 2002).
- L'ICD libéré de l'apoER2 se transloque vers le noyau et réprime la transcription Reelin (ApoER2 processing by presenilin-1 modulates reelin expression 2014 et RP8-Reelin-Regulated Neuronal Enhancer Signature Underlying Learning and Memory Formation 2015).
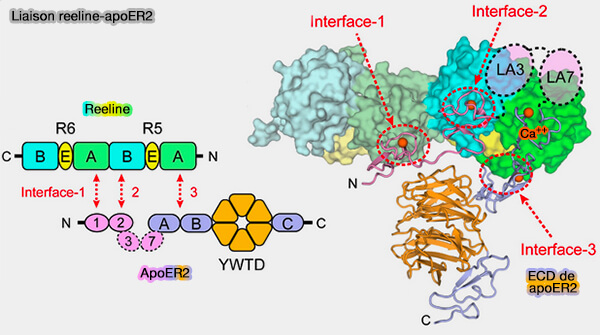
(Figure : vetopsy.fr d'après Hirai et coll)
Signalisation reeline non neuronale
L'ApoER2 et VLDLR interagissent avec bien d'autres récepteurs.
1. Par exemple, l'ApoER2 peut interagir avec VEGFR2 (Vascular Endothelial Growth Factor Receptor 2) pour faciliter la vascularisation dans le cortex en développement.
2. L'ApoER2 est aussi impliqué dans la voie de signalisation Wnt, et en particulier dans le métabolisme osseux ( LRP et tissu osseux).
3. La signalisation non neuronale Reelin pourrait également être impliquée dans le développement d'autres organes (Nonneuronal roles for the reelin signaling pathway 2016).
Métabolisme des lipides par VLDLR
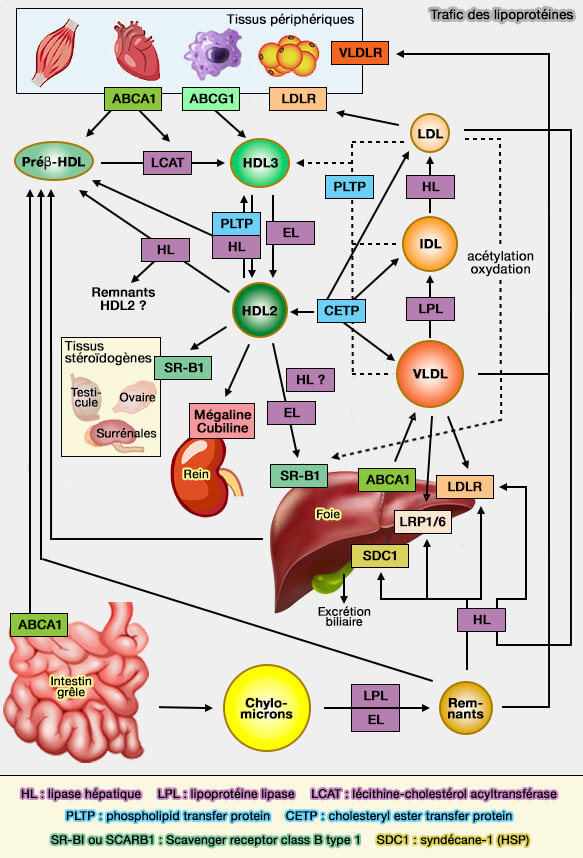
(Figure : vetopsy.fr)
VLDLR joue un rôle important dans :
- l'absorption du cholestérol,
- le métabolisme des TRL (lipoprotéines riches en triglycérides) contenant l'apoE.
Le VLDLR entre en interavtion avec les VLDL (lipoprotéines de très basse densité) et les IDL (lipoprotéines de densité intermédiaire), mais pas les LDL (lipoprotéines de basse densité) qui ne contiennent que l'apoB-100.
(Figure : vetopsy.fr d'après Dlugosz et Nimpf)
1. Dans les tissus périphériques, VLDLR agit dans l'endothélium pour améliorer l'hydrolyse dépendante de la LPL (LipoProtéine Lipase) des triglycérides (TG) et joue un rôle dans la clairance postprandiale des chylomicrons et des VLDL par leur absorption (The VLDL receptor plays a major role in chylomicron metabolism by enhancing LPL-mediated triglyceride hydrolysis 2004).
- Les souris Vldlr-/- présentent une diminution de l'expression expression de LPL, ce qui provoque une augmentation des TG plasmatiques.
- Les souris Vldlr-/- présentent des niveaux de TG plus élevés de 250% et une absorption de chylomicrons inférieure de 60% par rapport à celles du type sauvage [3].

VLDLR permet ainsi d'augmenter la production d'acides gras libres.
2. Il semblerait, d'après des études récentes, que l'équilibre approprié des niveaux de protéines VLDLR dans le système nerveux central (SNC) est important pour le maintien de l'homéostasie énergétique du corps entier (IDOL regulates systemic energy balance through control of neuronal VLDLR expression 2019).
a. Le rôle de l'hypothalamus est particulièrement important (Neural Control of Energy Balance: Translating Circuits to Therapies 2016), bien que l'équilibre énergétique soit maintenu par plusieurs régions du cerveau (Identification of a Brainstem Circuit Controlling Feeding 2017), en agissant :
- sur le système nerveux sympathique, pour son influence sur le tissu adipeux,
- sur les actions métaboliques de la leptine, du FGF21, des agonistes PPAR-γ (Peroxisome Proliferator-Activated Receptor γ) et de l'hormone thyroïdienne.
b. IDOL (inducible degrader of the LDLR) joue un rôle essentiel dans la dégradation des LDLR, des apoER2 et des VLDLR. Toutefois, la conséquence d'une carence en IDOL est une augmentation, plutôt qu'une diminution, des niveaux de VLDLR.
Cette région contribue probablement aux effets métaboliques d'IDOL par la détection des acides gras libres par l'hypothalamus qui est un modulateur bien établi de l'activité neuronale, affectant le métabolisme périphérique (A fatty acid-dependent hypothalamic–DVC neurocircuitry that regulates hepatic secretion of triglyceride-rich lipoproteins 2015).
Des études ultérieures sont nécessaires pour définir précisément comment l'activité IDOL dans le cerveau transmet un signal dépendant du VLDLR à la périphérie pour réguler l'équilibre énergétique et quelles populations neuronales spécifiques sont impliquées.
Retour aux LDLR
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesFormation des chylomicronsHDL et remodelageTrafic des lipoprotéinesRécepteurs des lipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation