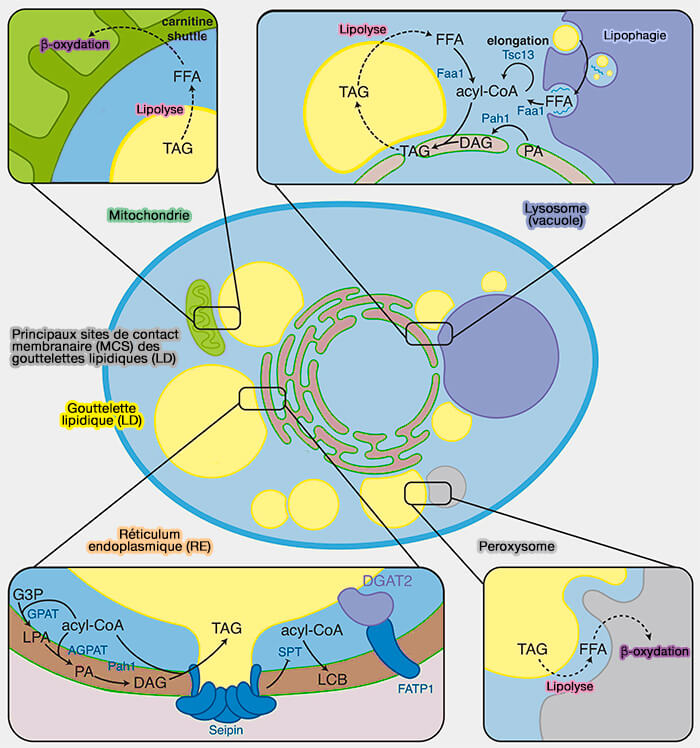
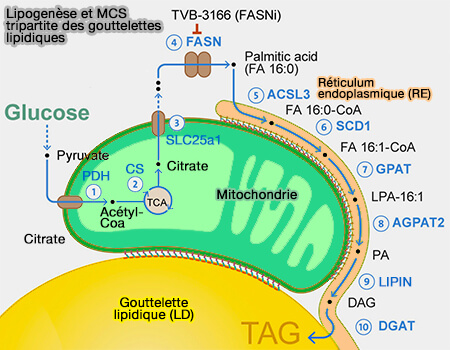
Remarque : de nombreux MCS sont tripartites, i.e. entre plusieurs organites, comme par exemple les MCS mitochondries-RE-LD pour le stockage des triglycérides (TG).
Principaux sites de contact membranaire (MCS) des gouttelettes lipidiques (LD)
(Figure : vetopsy.fr d'après Renne et Hariri)
L’activation de la réponse protéique déployée et le stress du RE peuvent stimuler la biogenèse des LD comme moyen de stocker les lipides tandis que les voies de synthèse des lipides dans le RE sont suspendues.
Ces MCS font partie d'un ensemble plus vaste appelé MCS tripartite : RE-LD-mitochondries.
Transfert des lipides
Les protéines de transport des lipides au niveau des contacts membranaires, où les organites sont étroitement liées, sont essentielles à la redistribution des lipides du réticulum endoplasmique (RE), où ils sont fabriqués, vers d'autres membranes cellulaires.
Un tel transfert médié par les protéines est particulièrement important pour maintenir les organites déconnectés des voies de sécrétion, comme les mitochondries.
MIGA2 dans les MCS
(Figure : vetopsy.fr d'après Freyre et col)
4. La manipulation de MIGA2 provoquent les conséquences suivantes.
La surexpression de MIGA2 dans les adipocytes entraîne une formation accrue de MCS LD-mitochondries.
Les adipocytes MIGA2-KO montrent une diminution de la différenciation adipocytaire et de la synthèse des triglycérides (TG) et la présence d'un grand nombre de petites LD.
Ce trafic se déroule au niveau des MCS LD-mitochondrie qui permettraient un passage rapide des lipides lors d'exercice par exemple, mais aussi pour prévenir la lipotoxicité.
Plusieurs protéines ont été impliquées dans la formation de ces MCS LD-mitochondrie.
La privation de glucose, une condition qui stimule l'oxydation des FA, favorise la co-immunoprécipitation de SNAP23, VAMP4 et ACSL1 dans les hépatocytes.
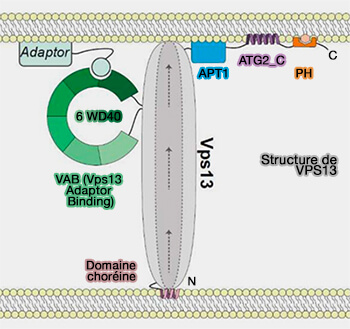
VPS13D
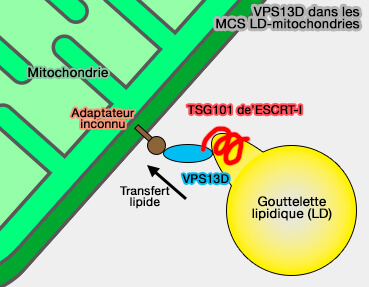
VPS13D, de la famille des VPS13 (Vacuolar Protein Sorting 13) transfère les FA des LD aux mitochondries au niveau des MCS entre ces organites.
1. Sa structure est composée de trois régions.
Sa région N-terminale est responsable du ciblage mitochondrial.
Un LTD (Lipid Transfer Domain) supposé à son extrémité N-terminale se lie aux FA pour le transfert.
Son domaine VAB (Vps13 Adaptor Binding) interagit avec TSG101/Vps23, une protéine de la tige de ESCRT-I pour le recrutement de TSG101 dans les MCS LD-mitochondries.
De plus, la localisation du domaine VAB et de TSG101/Vps23 entraîne la formation d'une structure resserrée ou tubulaire à la surface des LD.
Une suppression de VPS13D ou TSG101 entraîne une réduction significative du transfert de FA.
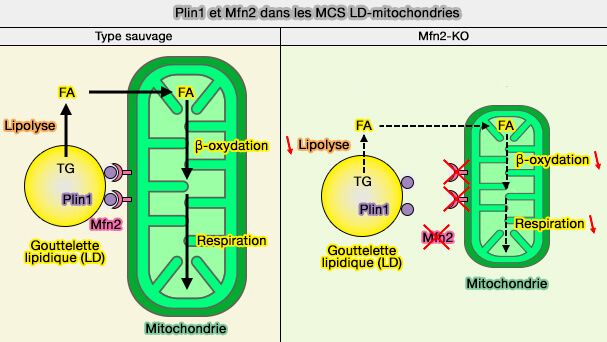
L'augmentation des contacts LD-mitochondries par MFN2-PLIN1 facilite le couplage de l'hydrolyse des triglycérides (TG) avec l'oxydation des FA lors de l'exposition du tissu adipeux brun au froid.
Rôle dans l'expansion des LD
Les MCS LD-mitochondrie peuvent également fonctionner dans l’expansion de LD dans des conditions favorisant le stockage des lipides.
VPS13D dans les MCS LD-RE
(Figure : vetopsy.fr d'après Benador et col)
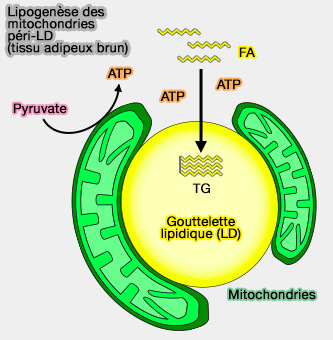
Ainsi, la formation des MCS LD-mitochondrie est associée au stockage des lipides et à la génération d'énergie par oxydation du glucose et non des FA, i.e. en fournissant de l'ATP pour la synthèse de l'acyl-CoA pendant la production de TG.
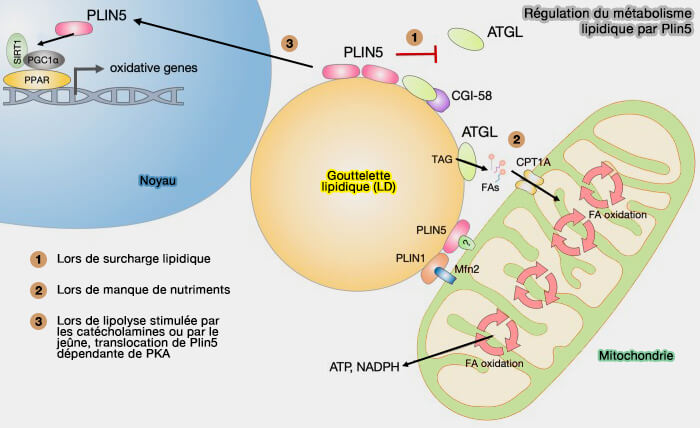
a. Les interactions de Plin5 facilitent la canalisation efficace des FA des cLD vers les mitochondries contrôlée par la signalisation PKA et la modulation ultérieure des interactions ATGL et CGI-58/ABHD5 ( Plin et lipolyse).
Régulation du métabolisme lipidique par Plin5
(Figure : vetopsy.fr d'après Jarc et Petan)
b. Toutefois, dans les muscles squelettiques humains, PLIN2 et PLIN5 se localisent dans les LD sur des sites distincts, avec une abondance de PLIN5 sur les sites d'attache LD-mitochondries.
c. On ignore si PLIN5 intervient directement dans l'attachement des gouttelettes lipidiques et des mitochondries par le biais d'une association avec une protéine mitochondriale spécifique ou avec la membrane mitochondriale.
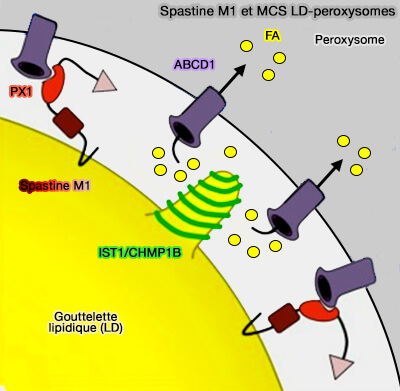
a. Elle forme, via sa région d'interaction avec les peroxysomes (PXI) qui se trouve au milieu de la protéine, un complexe d'attache avec le transporteur ABC, ABCD1 peroxysomal pour favoriser la formation de contact LD-peroxysome.
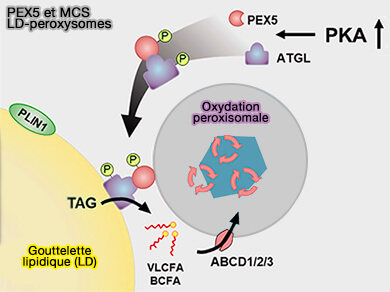
PEX5 et MCS LD-peroxysomes
(Figure : vetopsy.fr d'après Han et col)
b. Elle recrute les protéines d'ESCRT-III,IST1 et CHMP1B façonnant la membrane dans les LD via son domaine MIT pour faciliter le trafic d'acides gras (FA) du LD vers le peroxysome, éventuellement via des modifications dépendantes de IST1 et CHMP1B dans la morphologie de la membrane LD.
La perturbation de ce complexe d'attache modifie aussi les contacts peroxysome-RE et empêche la croissance de la membrane peroxysomale, ce qui suggère que ce site de contact est nécessaire pour transférer les lipides du RE vers les peroxysomes.







 MIGA1/2 et fusion mitochondriale),
MIGA1/2 et fusion mitochondriale),