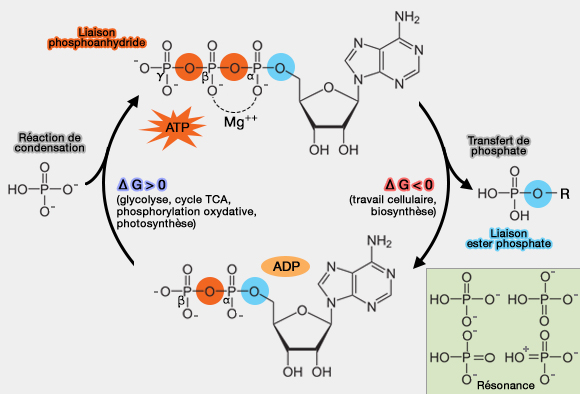
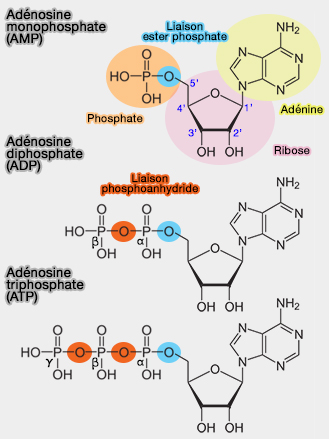
L’ATP (adénosine triphosphate) est un composé " riche en énergie " qui fournit l'énergie nécessaire à de nombreux processus biologiques grâce à son hydrolyse, mais a aussi d'autres fonctions.
Les composés " riches en énergie " sont des composés chimiques comportant une liaison chimique très facilement hydrolysable, et dont l'hydrolyse est accompagnée d'une très forte diminution d'énergie libre.
1. La valeur admise actuellement pour $\ce{\Delta G'0}$ (énergie libre ou énergie de Gibbs) lors de l'hydrolyse de la liaison phosphoanhydride terminale est d'environ 7,3 kcal.mole-1 ou 30,5 kJ.mole-1.
Il est très vraisemblable que cette valeur soit différente, sans doute plus élevée.
Elle dépend des concentrations locales en ATP, en ADP, en phosphate et en Mg++, et du pH.
Plus les concentrations des différents composants sont en équilibre, plus grande sera la valeur absolue du changement d'énergie libre de Gibbs.
2. Dans les conditions standard, i.e. les concentrations en ATP, ADP et Pi étant égales à 1M, la concentration en eau à 55M, la valeur de $\ce{\Delta G'0}$ est comprise entre -28 et -34 kJ / mol.
Cette réaction est catalysée par les ATPases , qui sont classées en plusieurs types, étudiées dans des chapitres spécifiques.
1. Par suite de la présence de charges négatives partielles des oxygènes des liaisons $\ce{P=0}$, les phosphores de la chaîne acquièrent une charge partielle positive, qui tend à entraîner une répulsion entre les atomes de phosphore, i.e. cela favorise l'hydrolyse de l'ATP.
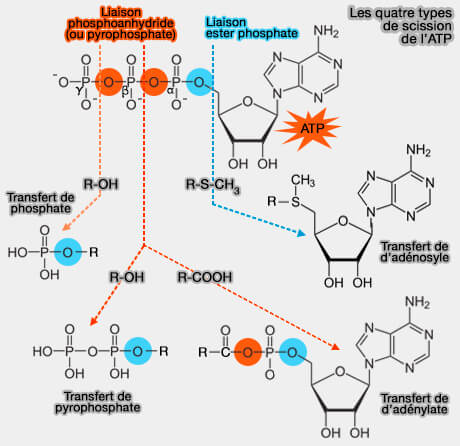
Les quatre types de scission de l'ATP
(Figure : vetopsy.fr)
a. À pH 7, les 3 restes phosphoriques sont ionisés, et l'ATP possède donc 4 charges négatives qui se repoussent fortement.
Ces groupes phosphate ont une stabilisation par résonance limitée car les électrons sont en partie entravés dans les liaisons entre phosphates.
La résonance est un concept qui explique comment une molécule peut répartir ses électrons sur plusieurs atomes pour être plus stable.
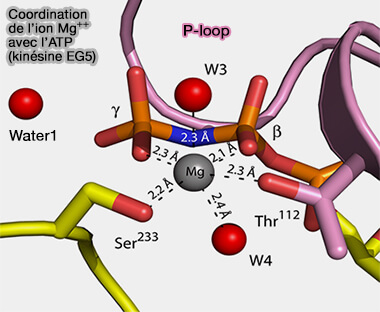
a. Les ions Mg++ permettent de neutraliser partiellement les charges négatives portées par les groupements phosphate de l’ATP, i.e. le complexe ATP/Mg++ est thermodynamiquement plus stable que l'ATP libre.
Coordination de Mg++ avec l'ATP
(Figure : vetopsy.fr d'après Parke et coll)
participer à l'hydrolyse en facilitant la rupture de la liaison phosphate par l'activation d'une molécule d’eau en la polarisant, facilitant ainsi son attaque nucléophile sur le groupe γ-phosphate,
stabiliser l’état de transition, i.e. conformation intermédiaire instable de l’ATP au moment de la rupture de la liaison au cours de la réaction enzymatique.
3. Après hydrolyse, les produits de réaction, ADP et phosphates, sont tous deux chargés négativement, i.e. n'ont pas tendance à se recombiner.
a. Le phosphate a plusieurs formes de résonance, i.e. les liaisons P=O et P–O- peuvent se déplacer grâce aux doublets libres des oxygènes, ce qui répartit la charge négative sur plusieurs atomes d’oxygène.
On dit qu'il y a délocalisation des électrons, et plus il y a de formes de résonance, plus l’ion est stable.
Pi ($\ce{HPO4^-- }$) a 4 formes de résonance à pH 7, qu'on peut simplifier en attribuant la double liaison à chaque oxygène à tour de rôle, mais en réalité, le phosphate est un hybride de ces quatre formes, ce qui signifie que la charge négative n’est pas localisée sur un seul oxygène, mais répartie équitablement entre tous.
Donc, la rupture de la liaison qui libère ce Pi est plus favorable thermodynamiquement, ce qui rend la réaction plus exergonique (ΔG°' très négatif), i.e. ADP + Pi est plus stable que l'ATP.
La stabilité de Pi est un facteur majeur qui rend l’hydrolyse de l’ATP très favorable énergétiquement.
b. En outre, après hydrolyse, Pi et ADP présentent des charges négatives plus exposées, ce qui favorise leur hydratation par une enveloppe de molécules d’eau formant des liaisons hydrogène (solvatation).
Cette solvatation contribue à réduire les répulsions électrostatiques entre charges, ce qui les rend plus stables du point de vue thermodynamique.
Impact acido-basique de l’hydrolyse de l’ATP
En raison des propriétés acide-base de l'ATP, de l'ADP et du phosphate inorganique, l'hydrolyse de l'ATP a pour effet d'abaisser le pH du milieu réactionnel par la libération d'un proton.
Lors d’efforts intenses, i.e. exercice anaérobie, on assiste à une forte hydrolyse de l’ATP, et l'accumulation de lactate, car les besoins énergétiques de la contraction musculaire dépassent le plus souvent les capacités de la mitochondrie à produire de l'ATP par la phosphorylation oxydative.
Ce processus contribue à l'acidose lactique. une acidose lactique, i.e. baisse du pH intracellulaire
Chez l'homme, environ 60 % de l'énergie libérée par l'hydrolyse d'une mole d'ATP produisent de la chaleur métabolique plutôt que d'alimenter les réactions réelles, ce qui est essentiel du point de vue physiologique.
Cette dissipation thermique, bien que représentant une perte d’énergie, est en réalité essentielle à la survie.
1. L’énergie libérée sous forme de chaleur contribue à maintenir la température interne stable ( homéothermie).
Cela est vital pour que les enzymes et les réactions biochimiques fonctionnent dans des conditions optimales.
2. Le corps ne peut pas utiliser toute l’énergie chimique directement dans des réactions mécaniques ou chimiques.
Une partie de l’énergie est inévitablement perdue sous forme de chaleur à cause de l’inefficacité des processus biologiques.
3. La chaleur produite participe aussi à la régulation des activités métaboliques et à la thermorégulation, comme la transpiration ou le frisson.
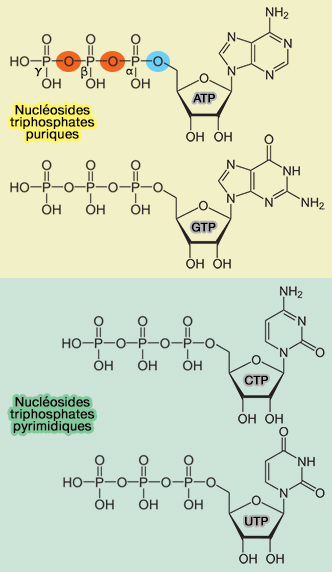
Production de l'ATP
On distingue deux sortes de phosphorylations de l'ADP en ATP ( production de l'ATP).
2. La phosphorylation oxydative est le processus permettant la phosphorylation de l'ADP en ATP grâce à l'énergie libérée par l'oxydation des donneurs d'électrons de la chaîne respiratoire.
Cette oxydation est rendue possible par les donneurs d'électrons NADH/FADH2 produits majoritairement par le cycle de Krebs.
Tous ces processus sont longuement étudiés dans des chapitres spécifiques.





 énergie libre ou énergie de Gibbs) lors de l'hydrolyse de la liaison phosphoanhydride terminale est d'environ 7,3 kcal.mole-1 ou 30,5 kJ.mole-1.
énergie libre ou énergie de Gibbs) lors de l'hydrolyse de la liaison phosphoanhydride terminale est d'environ 7,3 kcal.mole-1 ou 30,5 kJ.mole-1.