


Lipides
Acides gras
: catabolisme des acides gras
Bêta-oxydation (β-oxydation ou FAO)
Acides gras impairs et insaturés
β-oxydation peroxysomale
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Vue d'ensemble des lipides
- acides gras (FA)
- Classification des lipides
- Cholestérol
- Glycérides
- Trafic non vésiculaire des lipides et LTP
- Digestion et absorption entérocytaire des lipides
- Lipoprotéines
- Gouttelettes lipidiques (LD)
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La bêta-oxydation (β-oxydation ou FAO, fatty Acid β-Oxydation) se produit aussi sur les acides gras impairs saturés et sur les acides gras insaturés, mais également dans les peroxysomes des eucaryotes.
β-oxydation des autres acides gras dans les mitochondries
β-oxydation des acides gras impairs saturés
Les acides gras impairs sont rares chez les mammifères et sont retrouvés en particulier dans les triglycérides (Metabolism of Odd-numbered Fatty Acids and Even-numbered Fatty Acids in Mouse 2008).
1. Ces acides gras dont la synthèse a été décrite ( biosynthèse des acides gras impairs), subissent aussi la β-oxydation avec des produits finaux différents.
biosynthèse des acides gras impairs), subissent aussi la β-oxydation avec des produits finaux différents.
On aboutit au β-cétovaléryl-CoA qui est scindé par la β-cétothiolase (EC 2.3.1.16) en propionyl-CoA et acétyl-CoA.
$\ce{CH3-CH2-CO-CH2-CO-S-CoA}$ $\rightarrow$ $\ce{CH3-CH2-CO-S-CoA + CH2-CO-S-CoA}$
2. Le propionyl-CoA est transformé en succinyl-CoA qui peut alors entrer dans le cycle de Krebs.

(Figure : vetopsy.fr)
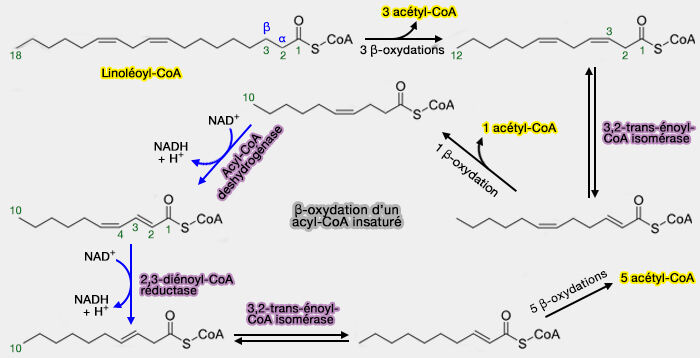
β-oxydation des acides gras insaturés
1. Le processus de la β-oxydation ne peut pas être sollicité lorsque une double-liaison cis inibe la formation de la liaison trans-Δ2, seul substrat possible pour la deuxième étape ( déshydrogénation de l'acyl-CoA).
Deux autres enzymes rentrent alors en ligne de compte. Le plus simple est de prendre un exemple pour comprendre le processus.
(Figure : vetopsy.fr)
2. Soit l'acide linoléique 18:2(n-6) ou 18:2(cis-Δ9,12).
a. Trois tours de β-oxydation conduisent au dodécénoyl-CoA en C12 avec une liaison cis-Δ3 qui ne peut être déhydrogéné par l'acyl CoA déshydrogénase.
b. Ce composé doit être isomérisé par la 3,2-trans-énoyl-CoA isomérase (EC 5.3.1.8) qui transforme la liaison cis-Δ3 en trans-Δ2.
c. Une nouvelle β-oxydation, suivi par une déhydrogénation aboutit à un décanoyl-CoA avec une liaison cis-Δ4 et trans-Δ2, i.e. configuration qui bloque cette fois l'hydratation par l'énoyl CoA hydratase.
d. La 2,4 diénoyl-CoA réductase (EC 1.3.1.34), enzyme à NADH qui peut convertir les liaisons trans-Δ2/trans-Δ4 ou trans-Δ2/cis-Δ4 en liaison trans-Δ3, forme en trans-Δ3-décanoyl CoA.
e. Ce composé est converti comme précédemment par la 3,2-énoyl CoA isomérase en trans-Δ2-décanoyl CoA qui peut alor subir cinq tours de β-oxydation pour abourir à l'acétyl-CoA.

La double liaison impaire (C3-C4) est transformée par l’isomérase en double liaison paire (C2-C3).
Les deux doubles liaisons paires (C2-C3 et C4-C5) sont converties par la réductase en une double liaison impaire (C3-C4).
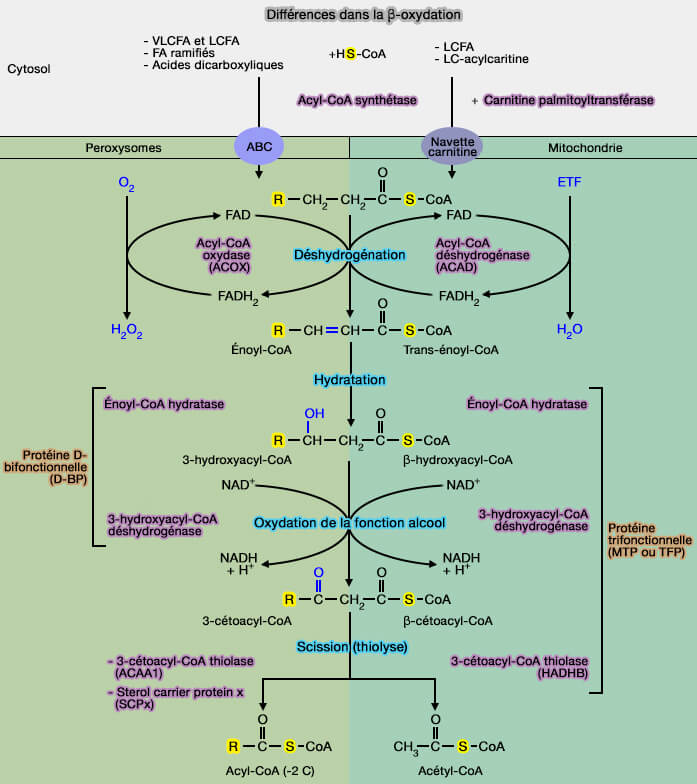
β-oxydation dans les peroxysomes
Dans les peroxysomes, ce sont avant tout les acides gras à longue et très longue chaîne > 22C, les acides gras ramifiés ainsi que les prostaglandines et les leucotriènes qui sont en partie dégradés et transféré dans les mitochondries pour achever leur dégradation complète en acétyl-CoA (Metabolic interactions between peroxisomes and mitochondria with a special focus on acylcarnitine metabolism 2020).
On note plusieurs différences avec la β-oxydation localisée des mitochondries.
1. Le transport des acides gras à longue chaîne (LCFA) dans les peroxysomes ne nécessite les carnitine palmytotransférase I et II (CPT1 et CPT2), mais des transporteur ABC, i.e. ABCD1, ABCD2 et ABCD3.
a. La mutation de ABCD1, localisé sur le chromosome X, provoque l'adrénoleucodystrophie liée à l'X (X-ALD), se traduit par une démyélinisation progressive du système nerveux central et une insuffisance surrénalienne par l'accumulation d'acides gras linéaires saturés à très longue chaîne, notamment l'acide cérotique (C26:0) dans le cerveau, la moelle épinière, les glandes surrénaliennes et les cellules de Leydig testiculaires ( Adrénoleucodystrophie).
b. ABCD1 forme avec la spastine M1 un complexe d'attache pour favoriser la formation de contact gouttelette lipidique (LD)-peroxysome.
(Figure : vetopsy.fr)
2. La première étape de cette dégradation, différente de celle de la voie mitochondriale, est réalisée par une enzyme spécifique, l'acyl-CoA oxydase (EC 1.3.3.6) qui catalyse la réaction :
$\ce{Acyl-CoA + O2}$ $\leftrightharpoons$ $\ce{Trans-Δ2-énoyl-CoA + H2O2}$
a. Dans la réaction de désydrogénation catalysée par l'acyl-CoA déshydrogénase, les électrons à haut potentiel sont récupérés sous forme de FADH2, permettant de libérer leur énergie métabolique sous forme d'ATP par la chaîne respiratoire, i.e. 1 FADH produisant 2 ATP ( bilan de la β-oxydation).
b. Dans les peroxysomes, $\ce{H2O2}$ est converti par la catalase, enzyme spécifique aux peroxysomes.
$\ce{2 H2O2}$ $\leftrightharpoons$ $\ce{2 H2O + O2}$
3. Dans les peroxysomes, l'énoyl-CoA hydratase et la 3-hydroxyacyl-CoA déshydrogénase, i.e. actives dans la 2ème et 3ème étape de la β-oxydation, forme un complexe enzymatique, i.e. protéine D-bifonctionnelle, ce qui diffère de la protéine trifonctionnelle mitochondriale (MTP ou TFP).
4. La 3-cétoacyl-CoA thiolase peroxysomale, i.e. ACAA1 (EC 2.3.1.155) distincte de celle des mitochondries, i.e. HADHB, active dans la scission, est très efficace que sur les acides gras à plus de huit atomes de carbone, d'où l'accumulation d'octanoyl-CoA, qui doit être oxydé dans les mitochondries.
Toutefois, SCPx (Sterol Carrier Protein x) a aussi un rôle de thiolase dans les peroxysomes.
(Figure : vetopsy.fr d'après Houten et coll)
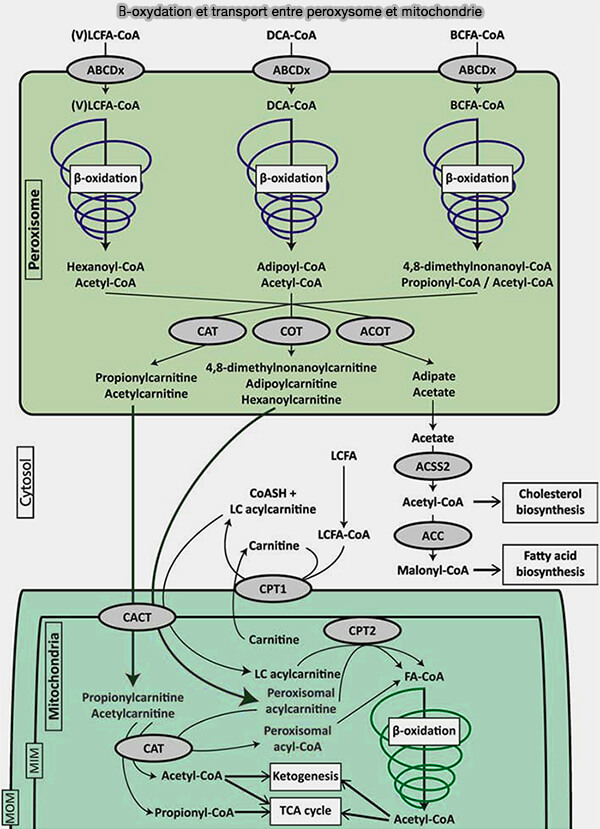
4. La dégradation des différents acides gras dans les péroxysomes aboutit :
- pour les acides gras à longue chaîne à l'octanoyl-CoA, i.e. C8, mais aussi en en hexanoyl-CoA, i.e C6 suivant certains auteurs,
- pour les acides dicarboxyliques-CoA en adipoyl-CoA,
- pour les acides gras ramifiés en 4,8-diméthylnonanoyl-CoA et propionyl-CoA,
- pour tous, on peut aussi arriver à de l'acétyl-CoA déjà dans les peroxysomes.
5. Ces acyl-CoA vont traverser la membrane des peroxysomes par des carnitine transférases différentes des carnitine palmitoyltransférases (CPT1 et 2) des mitochondries.
a. La carnitine octanoyltransférase (CROT ou COT), i.e. EC 2.3.1.137, spécifique des peroxysomes, intervient pour les octanoyl-CoA, mais aussi semble-t-il, pour les 4,8-diméthylnonanoyl-CoA et les adipoyl-CoA.
$\ce{Octanoyl-CoA + carnitine}$ $\leftrightharpoons$ $\ce{O-octanoyl-carnitine + CoA}$
b. La carnitine O-acétyltransférase (CRAT ou CAT), aussi présente dans les mitochondries, fait traverser les acétyl-CoA et propionyl-CoA .
$\ce{Octanoyl-CoA + carnitine}$ $\leftrightharpoons$ $\ce{O-octanoyl-carnitine + CoA}$
c. Les acyl-CoA thioestérases (ACOT), i.e. EC 3.1.2.-, peuvent aussi transformer les adipoyl-CoA et acétyl-CoA en adipates et les acétates qui traversent la membrane.
$\ce{Acétyl-CoA + H20}$ $\leftrightharpoons$ $\ce{Acétate + CoA}$
Remarque : certaines ACOT font partie des protéines STARD, protéines de transfert des lipides (LTP ou Lipid Tranfer Protein) impliquées dans leur trafic non vésiculaire, i.e. STARD14 (ou ACOT11) et STARD15 (ou ACOT12).
Retour à la β-oxydation
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesacides gras (FA)Synthèse des acides grasβ-oxydationCorps cétoniquesOxylipinesLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesGlycérophospholipidesSphingophospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation