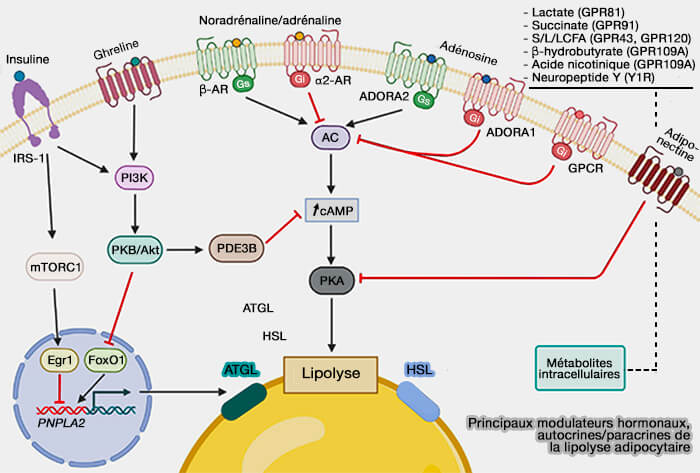
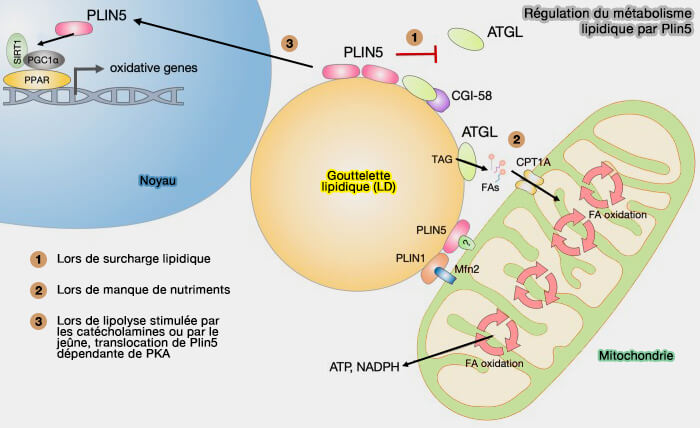
Cette fonction est facilitée par la réduction de l'activité de la PKA induite par l'insuline par plusieurs mécanisme lors de l'état postprandial ( régulation de la lipolyse adipocytaire).
3. Plin1 forme un échafaudage stratégique à la surface des cLD du tissu adipeux blanc (WAT) pour l'assemblage et le désassemblage coordonnés de complexes lipolytiques multiprotéiques lors de la phosphorylation ou de la déphosphorylation de Plin1a.
Dans la lipolyse, CGI-58/ABHD5 a un rôle essentiel qui est étudié dans un chapitre spécifique
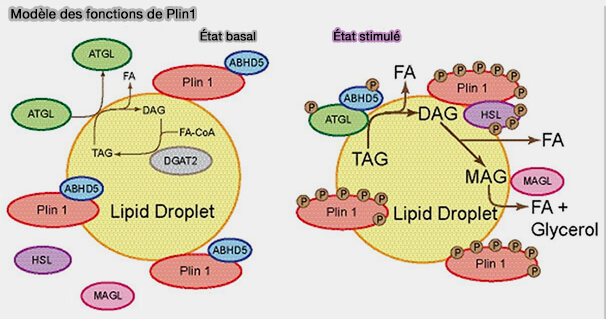
1. D'une part, l'extrémité C-terminale de Plin1a non phosphorylée interagit avec CGI-58/ABHD5 l'activateur de l'ATGL, limitant ainsi l'activité lipolytique cellulaire.
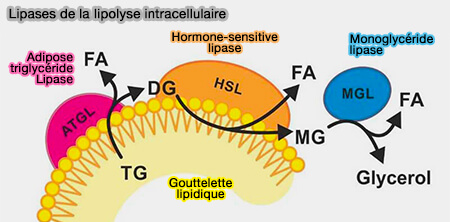
Dans des conditions basales, les lipases sont séquestrées des cLD et la lipolyse est supprimée.
Principaux modulateurs hormonaux, autocrines/paracrines de la lipolyse adipocytaire
(Figure : vetopsy.fr d'après Li et coll)
Dans ces conditions, Plin1, CGI-58/ABHD5 et ATGL, ainsi qu'HSL sont phosphorylées par la PKA avec pour conséquence une réorganisation de l'échafaudage des cLD qui est essentielle pour l'activation massive de la lipolyse, i.e. plus de 50 fois.
1. La phosphorylation de Plin1 libère CGI-58/ABHD5 phosphorylé, qui interagit avec l'ATGL phosphorylé pour coactiver l'hydrolyse des triglycérides (TG).
L'ATGL phosphorylée est fortement recrutée dans les gouttelettes lipidiques où elle se lie de manière indépendante de Plin1.
2. La HSL phosphorylée subit une translocation du cytoplasme vers la surface des gouttelettes lipidiques, où elle se lie à Plin1 phosphorylée et hydrolyse les diglycérides (DAG) produit par l'ATGL.
Remarque : actuellement, on ne sait pas si Plin1 contrôle ou non l'activité ou la localisation de MGL/MAGL (MonoacylGlycérol Lipase) dans les gouttelettes lipidiques, ou si la localisation de MGL est modifiée par la stimulation.
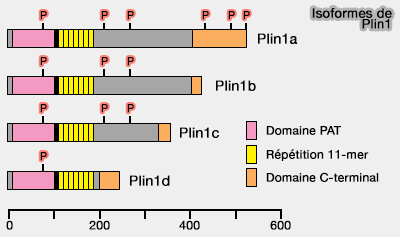
a. Plin1b et Plin1c ne possèdent pas les trois sites C-terminaux, ce qui peut expliquer leur faible protection contre la lipolyse et la mauvaise réponse à l'activation de la PKA.
b. Plin1a humain n'a pas de site 2, et des insertions de résidus par rapport à celui de la souris implique que les sites 5/6 correspondent aux Ser497/522.
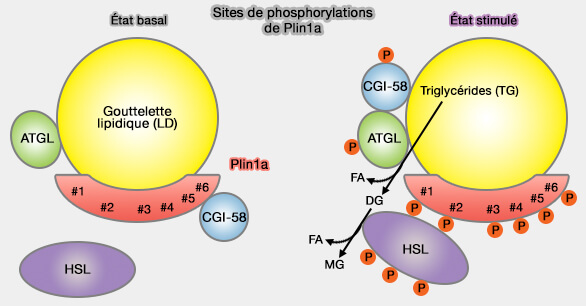
Sites de phosphorylations de Plin1a
(Figure : vetopsy.fr)
2. Les sites 5/6 C-terminaux de phosphorylation de la PKA sont essentiels.
a. La phosphorylation C-terminale de Plin1a perturbe l'interaction avec CGI-58/ABHD5.
Néanmoins, CGI-58 reste lié à la surface du CLD via son extrémité N-terminale riche en tryptophane, i.e. Trp-21, Trp-25 et Trp-29 ( plus haut).
b. La phosphorylation par PKA de CGI-58 améliore sa capacité à recruter, mobiliser et activer l'ATGL.
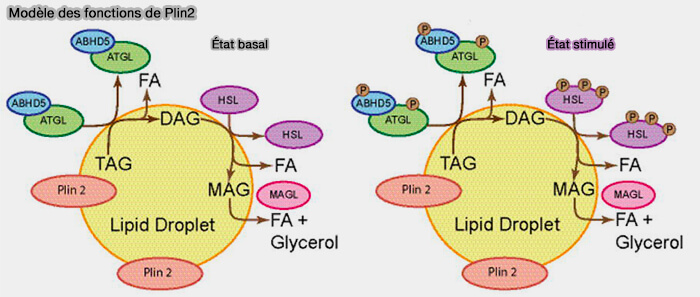
Plin2, est exprimée de manière ubiquitaire, est la principale périlipine associée aux gouttelettes lipidiques dans les cellules dans lesquelles Plin1 ou Plin5 sont réduites ou absentes.
La périlipine 2 n'atténue que modérément la lipolyse et n'est pas phosphorylée par la PKA.
La périlipine 2 se localise presque exclusivement dans les gouttelettes lipidiques et ne recrute pas de manière significative CGI-58/ABHD5, ATGL ou HSL dans les gouttelettes lipidiques.
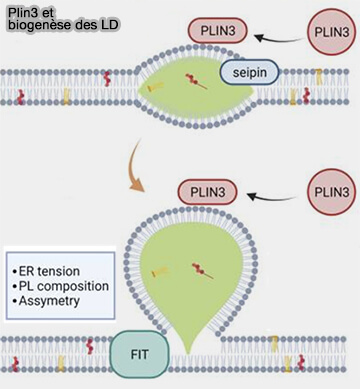
Plin3 et biogenèse des LD
(Figure : vetopsy.fr d'après Kaiser et coll)
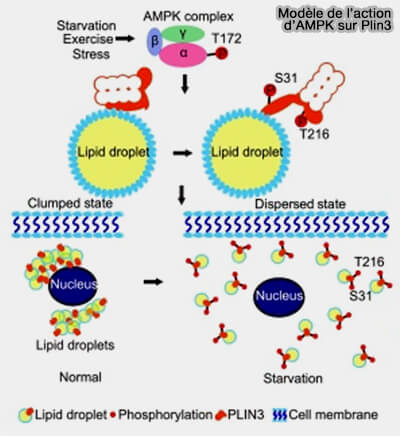
Elle n'est pas phosphorylée par la PKA et la lipolyse dans des conditions basales est approximativement comparable à la lipolyse lors d'activation de la PKA.
Plin3
Plin3 est exprimée de manière omniprésente dans les tissus et les lignées cellulaires en culture.
Plin4 est associée à de gouttelettes lipidiques naissantes minuscules d'adipocytes en culture et aux gouttelettes lipidiques du tissu adipeux blanc des tissus oxydatifs, i.e. cœur et muscle squelettique (Adipocyte Protein S3-12 Coats Nascent Lipid Droplets 2003).
Son inactivation serait associée à une réduction de la teneur en ARNm et en protéines de Plin5, en particulier dans le cœur, diminuant ainsi l’accumulation de lipides cardiaques chez la souris.
Toutefois, Plin4 et Plin5 sont localisés sur une région très proche du même chromosome et ce pourrait être un artefact.
2. Plin4 ne semble pas la cible de la phosphorylation, mais son étude est des plus limitée.
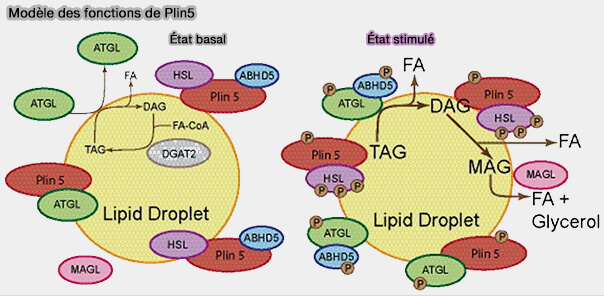
Plin5
Plin5 diminue l'hydrolyse des cLD dans les tissus oxydatifs, i.e. cœur, muscle squelettique et foie, avec Plin2, et tissu adipeux brun avec Plin1, mais le mécanisme est encore à clarifier.
1. Les expérimentations ont montré un processus un peu différent de celui de Plin1 ( cf. plus haut).
2. Plin5 est modélisé comme un échafaudage cLD formant une barrière lipolytique qui séquestre les substrats des cLD des lipases, i.e. en se liant, comme, Plin1, au CGI-58/ABHD5 et à HSL, mais aussi à l'ATGL.
La liaison à l'ATGL et à CGI-58/ABHD5 s'exclue mutuellement, i.e. les sites de liaison se chevauchent, empêchant les deux protéines de se lier à une seule molécule de Plin5.
c. Le site de liaison pour HSL est distinct et est localisé, comme Plin1, à l'extrémité N-terminale.




 régulation de la lipolyse adipocytaire).
régulation de la lipolyse adipocytaire).