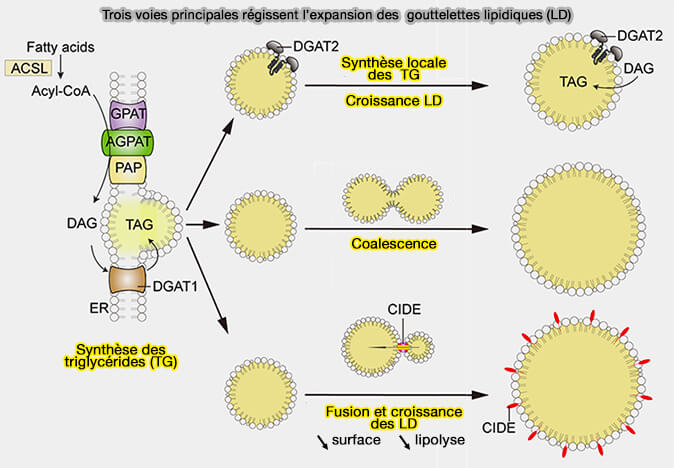
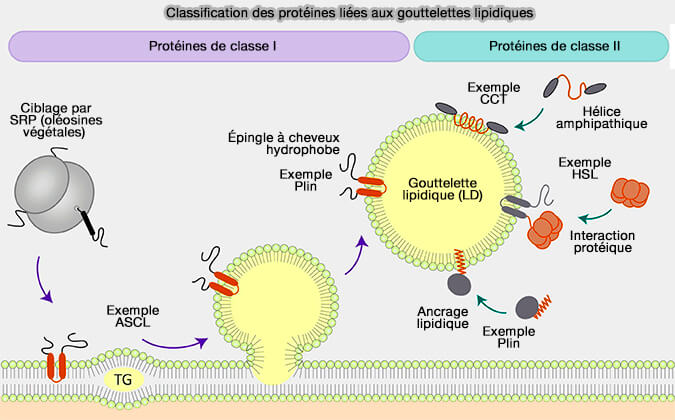
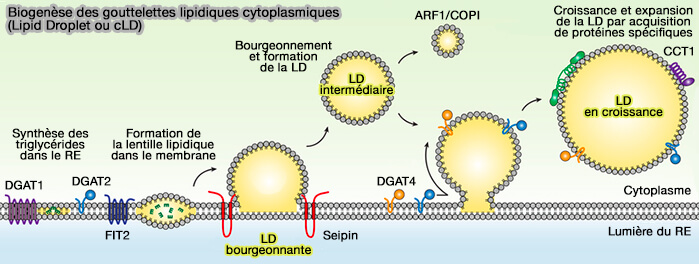
2. Pendant ces processus elles peuvent accroître leur volume, mais la taille des LD n’est pas uniquement fonction de la charge cellulaire en lipides neutres, i.e. triglycérides (TG) et esters de cholestérol (CE). Elle peut être régulée par :
b. Cette coalescence pourrait déjà intervenir dans la formation des lentilles lipidiques.
2. Le mûrissement (ripening) d'Ostwald favorise le transfert de lipides neutres depuis des gouttelettes lipidiques plus petites vers des gouttelettes lipidiques plus grosses et plus stables.
En effet, si de nombreuses LD sont nucléées parallèlement, il serait défavorable à toutes de croître (Lipid Droplet Nucleation 2023).
Si les LD ne sont pas stabilisés contre la maturation, l’état d’équilibre exige qu’une seule LD, la plus grande, croisse aux dépens des autres qui se résorbent.
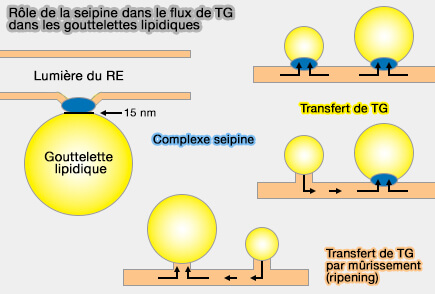
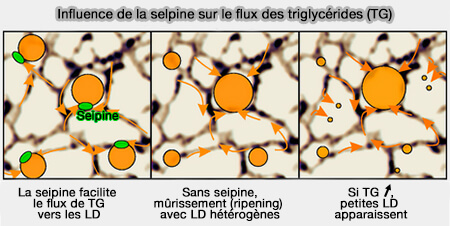
Influence de la seipine sur le flux de TG vers les LD
(Figure : vetopsy.fr d'après Salo et coll)
Les LD ne se développent pas normalement en l'absence de seipine.
Si la seipine est relocalisée dans le RE, la croissance de la LD a lieu sur ce site.
Si la nombre de seipines est réduit, la croissance des LD est préférentiellement ciblée sur ces sites.
Dans un réseau RE continu, les LD dépourvues de seipine rétrécissent, tandis que celles contenant de la seipine se développent.
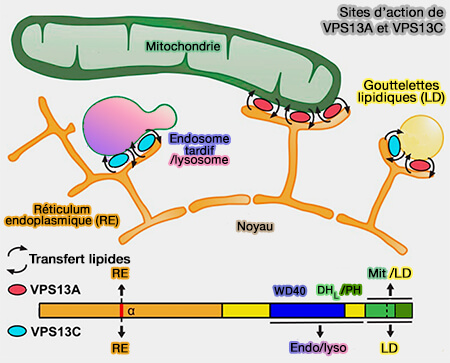
Outre la synthèse in situ et la diffusion, les phospholipides (PL) peuvent être délivrés directement du RE aux LD via des protéines de transfert lipidique opérant au niveau des sites de contact RE-LD ou mitochondries-LD.
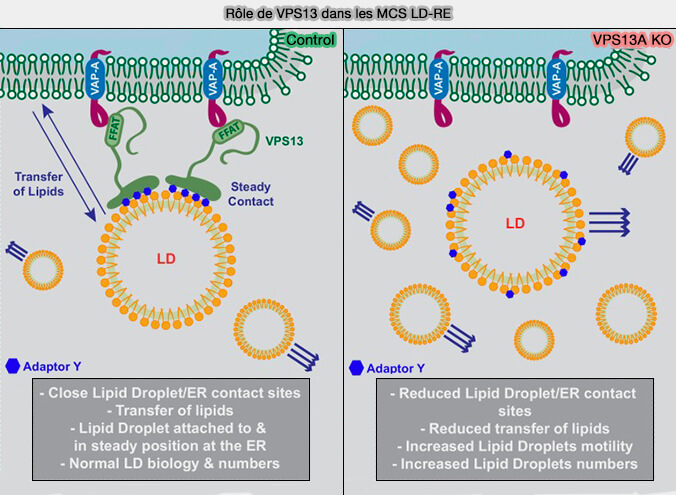
Rôles de VPS13A dans les MCS LD-RE
(Figure : vetopsy.fr d'après Yeshaw et coll)
2. D'autres protéines de transfert de lipides sont aussi engagés dans ce processus comme les membres de la famille ORP/Osh, i.e. Oxysterol-binding proteins(OSBP)-Related Proteins mammaliennes et OsBP-homolog de la levure.
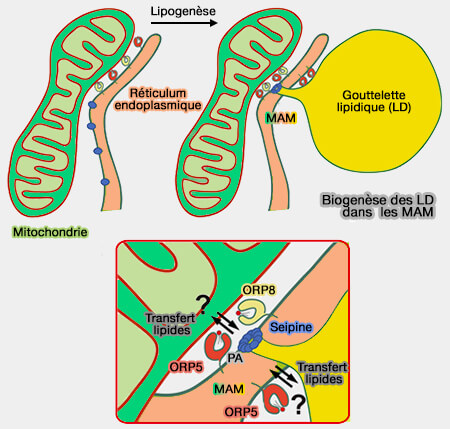
Biogenèse des LD dans les MAM
(Figure : vetopsy.fr d'après Guyard et coll)
Au cours de l'expansion des LD, la tension superficielle/linéaire de la monocouche LD doit être permissive et cette propriété est subordonnée à la composition et les défaut de compactage de lipides de la monocouche.
1. L'enrichissement en lipides formant des bicouches tels que la phosphatidylcholine (PC) à la surface des LD peut aider à stabiliser les LD en partie en minimisant les défauts de compactage.
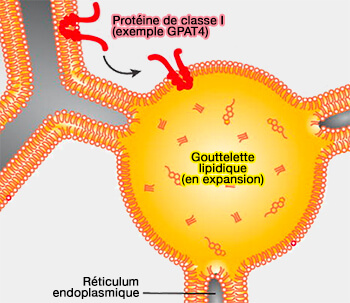
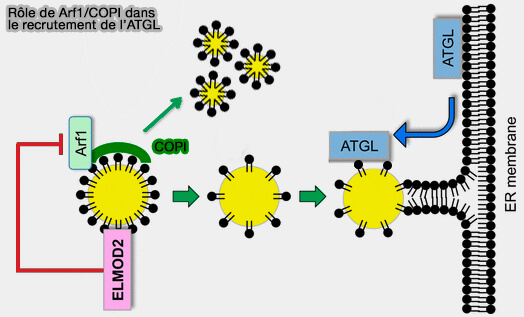
Rôle d'ARF1/COPI dans le recrutement de l'ATGL
(Figure : vetopsy.fr d'après Suzuki et coll)
a. Par ce processus, Arf1/COPI éliminerait les phospholipides (PL) des surfaces des LD.
Le bourgeonnement des nano-LD entraîne la réduction du tassement phospholipidique et l'augmentation de la tension superficielle de la surface de la LD qui induit des ponts membranaires entre les LD et la membrane du RE à travers lesquels les protéines de classe I (ERTOLD) comme l'ATGL peuvent diffuser latéralement vers les LD.
Les cellules dépourvues de la fonction Arf1/COPI contiennent plus de phospholipides sur les LD, ce qui entraîne une diminution de la tension superficielle des LD et une incapacité à former des ponts vers le RE.
Il joue un rôle dans la fusion et la croissance des gouttelettes lipidiques.
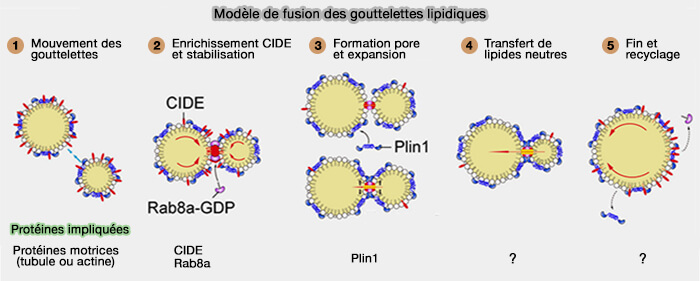
2. Les oligomères CIDE, en plus d'attacher deux LD, formeraient des pores ou des canaux pour le passage des triglycérides (TG) entre les LD, invariablement de la plus petite LD à la plus grosse grâce aux différences de pression interne (mûrissement (ripening) d'Ostwald), aidés en cela par PLIN1 et Rab8a.
Les CIDE sont étudiées dans un chapitre spécifique.
Modèle de fusion des gouttelettes lipidique
(Figure : vetopsy.fr d'après Chen et coll)




 ponts membranaires).
ponts membranaires).