Dans les adipocytes, les LD sont plus grosses et remplissent la grande partie de la cellule.
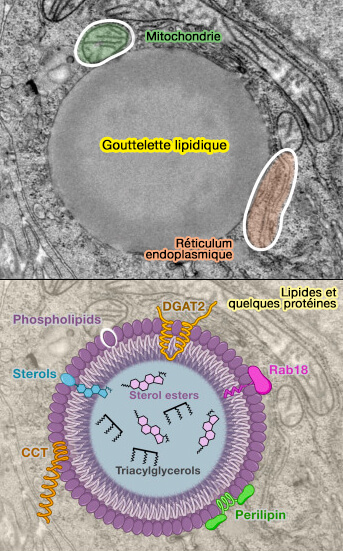
Structure simplifiée des LD
(Figure : vetopsy.fr d'après Farese et coll)
Dans d'autres cellules, elles sont plus petites et souvent produites dans certaines conditions.
2. Les LD sont composées d'une monocouche dont la composition est identique à celle de la membrane du réticulum endoplasmique (RE), mais qui lui confère des propriétés spécifiques.
On n'a pas encore découvert de machinerie protéique qui intervient dans l'insertion directe de protéines dans la surface des LD ou de marqueurs lipidiques comme les différents phosphoinositides pour le ciblage des LD.
Particularités de la monocouche
Composition de la monocouche
La composition de la monocouche varie en fonctions de nombreux facteurs et est essentielle :
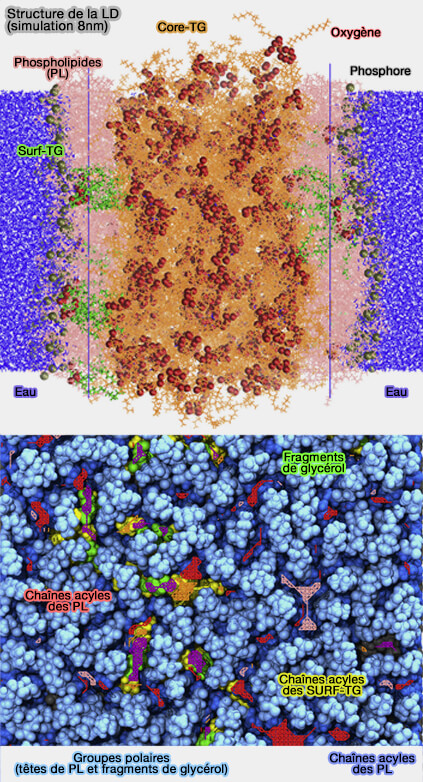
Membrane simulée des LD
(Figure : vetopsy.fr d'après Kim et coll)
Les résultats des simulations de l'article précédent suggèrent que 5 à 8 % de la surface de la LD est occupée par des molécules de triglycérides (TG), un nombre qui dépasse la solubilité maximale rapportée pour les TG dans les bicouches phospholipidiques (2,8 %).
1. On trouve deux classes distinctes de molécules TG qui interagissent avec la monocouche des LD.
a. Les SURF-TG, à la surface de la monocouche, sont ordonnées comme les phospholipides (PL) avec le fragment glycérol exposé à l'eau, créant une quantité importante de défauts d'emballage chimiquement uniques et les chaînes acyles étendues vers le centre de la LD.
b. Les CORE-TG, i.e. TG qui s'intercalent uniquement dans la région de la queue des PL, sont désordonnés et augmentent la quantité de défauts de compactage et de l'ordonnancement de la queue des PL.
c. Le degré d'interdigitation provoqué par CORE-TG est stable et détermine la largeur du chevauchement TG-PL, tandis que celui provoqué par SURF-TG fluctue et est fortement corrélé à la surface par PL ou à l'expansion de la monocouche.
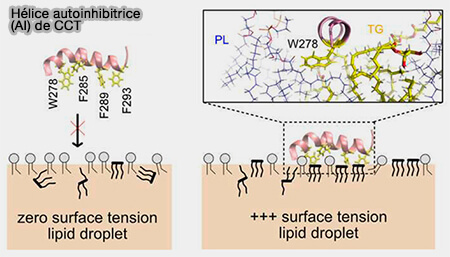
Lorsque l’offre de PL est limitée, SURF-TG peut réduire la tension superficielle en se comportant comme un composant secondaire de la membrane.
Ces enzymes expliquent les alternances de taille des LD selon la lipogenèse ou la lipolyse, i.e. lipolyse enzymatique neutre, mais aussi par une forme sélective d'autophagie (lipophagie) lors des cycles métaboliques.
Remarque : toutefois, en raison des difficultés liées à l’obtention de préparations pures de LD, combinées à la sensibilité croissante de la spectrométrie de masse, de nombreuses listes de protéomes de LD contiennent des contaminants protéiques, ce qui explique les différences selon les publications.
4. Les protéines sont identiques dans les cellules, excepté dans les adipocytes et les hépatocytes, cellules hautement spécialisées pour stocker les lipides neutres et qui possèdent des protéines spécifiques pour réguler certains processus des LD.
La périlipine1 (Plin1) se trouve exprimée uniquement dans les adipocytes, où elle joue un rôle clé dans la régulation de la lipolyse induite par le jeûne.
Ciblage des protéines sur les gouttelettes lipidiques
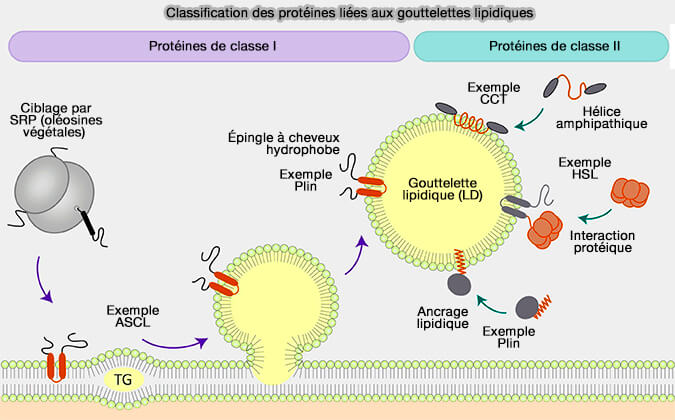
Classification des protéines liées aux gouttelettes lipidiques
(Figure : vetopsy.fr d'aprèsDhiman et coll)
1. Les contraintes biophysiques des LD, i.e. noyau lipidique neutre hydrophobe et monocouche de phospholipides (PL), est un environnement énergétiquement défavorable pour les domaines protéiques hydrophiles, i.e. le noyau lipidique du LD ne contient aucune protéine.
L'architecture de la monocouche LD, obligent les protéines LD à adopter des conformations évitant d'exposer les domaines hydrophiles au noyau lipidique neutre de la LD. Pour cette raison, les protéines LD intégrales sont des protéines monotopiques qui s'insèrent dans la monocouche LD et orientent leurs domaines solubles vers le cytosol.
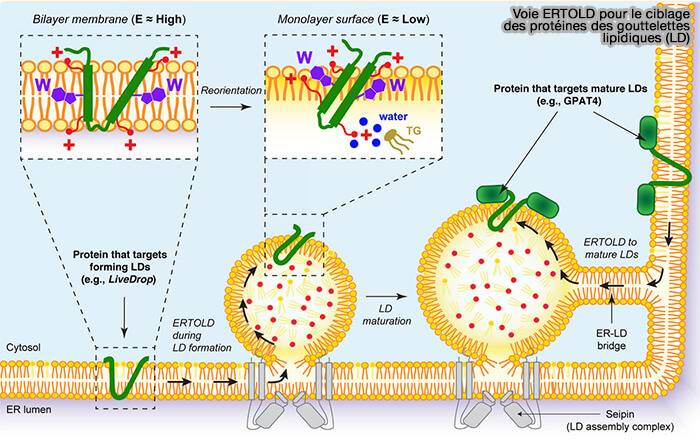
La classe I est composée de protéines à double localisation membranaire, i.e. du réticulum endoplasmique (RE) ou LD via des domaines hydrophobes, en général en épingle à cheveu (hairpin en anglais).
Bien que la structure de ces séquences hydrophobes ne soit pas connue, elles semblent dépourvues de domaines luminaux, ce qui leur permet de s'intégrer dans les deux membranes.
Les deux extrémités C- et N-terminales sont localisées dans le cytosol.
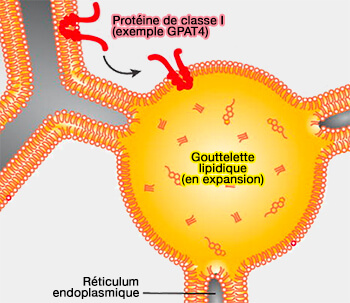
Protéines de classe I : GPAT4
(Figure : vetopsy.fr d'après Kory et coll)soit pendant la formation des LD (iLD ou initial LD) et subissent une translocation vers la LD en formation, comme par exemple l'ACSL (long-chain-fatty-acid-CoA ligase) qui active les chaîne d'acyl gras (cf. figure ci-dessus),
2. Le mécanisme d'élimination de ces protéines de la surface des LD reste encore à préciser, mais devrait être régulé par la charge lipidique de la LD.
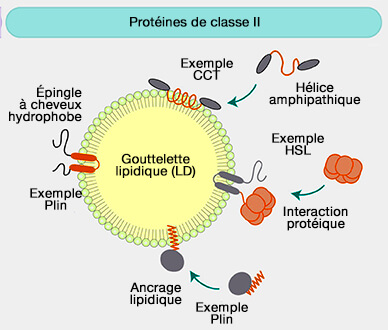
Classe II (CYTOLD)
1. La classe II comprend des protéines cytosoliques qui subissent une translocation, i.e. elles sont appelées aussi CYTOLD (Cytoplasme to LD), et s'attachent à la monocouche par plusieurs mécanismes.
Protéines de classe I : GPAT4
(Figure : vetopsy.fr d'après Dhiman et coll)
b.
Les périlipines (Plin) s'ancrent par leurs nombreuses hélices hydrophobes (répétitions 11-mer), i.e. différentes des AH car tous leurs résidus sont hydrophobes, et des domaines hydrophobes comme leur domaine C-terminal,
c. D'autres protéines comme Rab18 ou ELMOD2, une Arf-GAP non canonique, sont des protéine TA (Tail-Anchored, i.e. ancrée par la queue) qui s'accrochent par ancrage lipidiques.
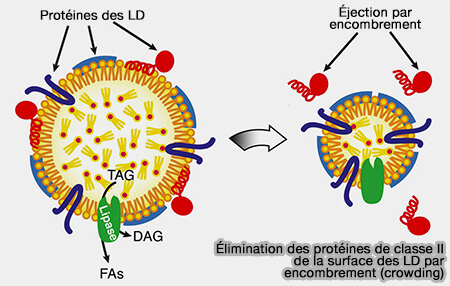
Élimination des protéines de classe II par encombrement
(Figure : vetopsy.fr d'après Kory et coll)
Lors de la lipolyse, la perte de volume des LD diminue leur surface, comprime les protéines les unes contre les autres et augmente la pression superficielle, i.e. les protéines sont éjectées dans un processus similaire aux apolipoprotéines échangeables.
En outre, la perte de surface limite les sites de liaison disponibles, empêchant ainsi les protéines déplacées de se lier à nouveau aux LD.
b. Les modifications post-traductionnelles comme les cycles de phosphorylation et déphosphorylation qui, par exemple, modifie l'affinité des protéines pour la surface.





 élimination des CYTOLD).
élimination des CYTOLD).