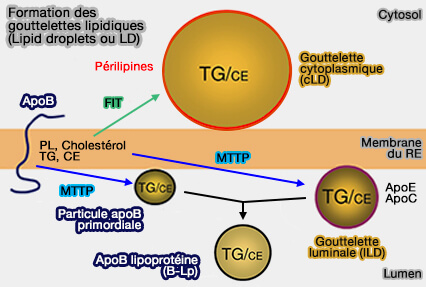
soit dans le cytoplasme, pour former des gouttelettes lipidiques cytoplasmiques (cLD) libres,
soit dans la lumière du RE, pour former gouttelettes lipidiques luminales (lLD).
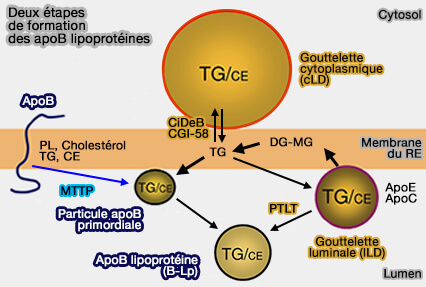
La biogenèse des gouttelettes lipidiques nucléaires (lLD) s'effectuerait selon deux mécanismes.
Remarque : les gouttelettes lipidiques cytoplasmiques (cLD) ne peuvent pas forcément dire qu'elles sont libres dans le cytosol, car elles sont, dans leur grande majorité (85%), attachées au réticulum endoplasmique (RE).
La kinésine qui transporte les cLD le long des microtubules, serait principalement impliquée dans l'apport de TG aux B-lps lors de l'assemblage.
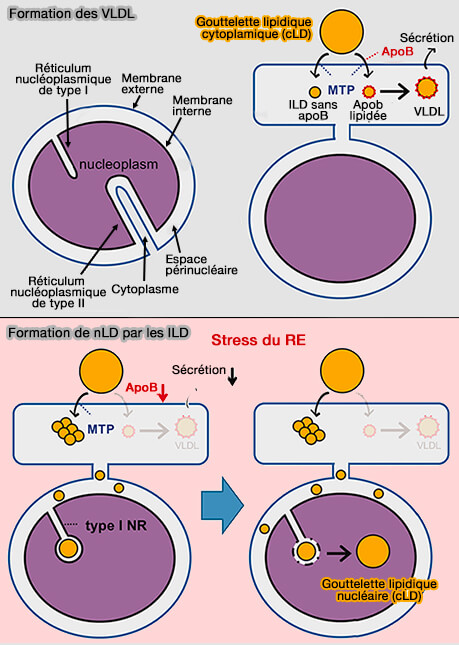
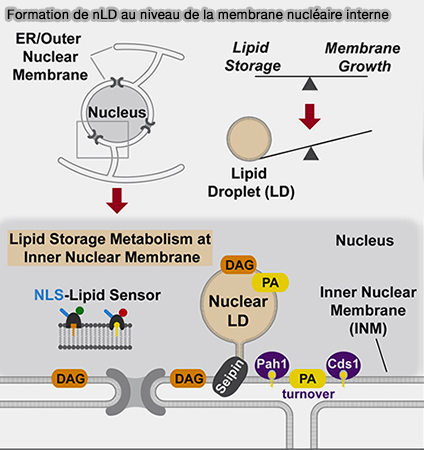
Formation des nLD à partir des lLD
(Figure : vetopsy.fr d'après Sołtysik et coll)
La kinésine peut transporter les cLD vers le RE, et ces cLD peuvent fournir des précurseurs lipidiques pour l'assemblage des VLDL lors de satiété.
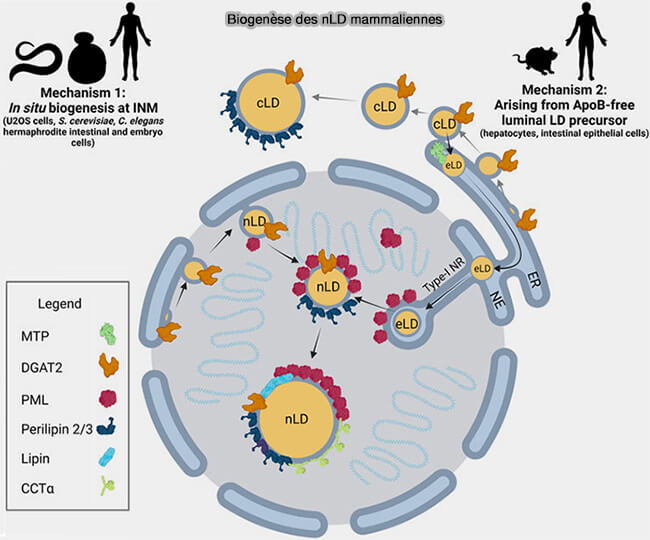
Les gouttelettes lipidiques nucleaires (nLD) ne sont pas des cLD piégées dans le noyau, mais bien de véritables organites dotés de leurs propres caractéristiques et leur biogenèse s'effectuerait selon deux processus.
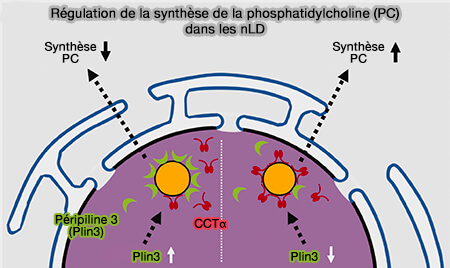
Régulation de la synthèse de PC dans les nLD
(Figure : vetopsy.fr d'après Sołtysik et coll)
La périlipine-3 (Plin3) entre en compétition avec le CCTα dans la liaison nucléaire de la LD et semble fonctionner comme un régulateur du CCTα.
2. Les nLD semblent réguler la structure de la chromatine, l'expression des gènes et la signalisation cellulaire, i.e. le sujet est complexe et nous entraînerait trop loin.
Par exemple, elles synthétiseraient des facteurs de transcription, tels que Opi1, à proximité immédiate de leurs gènes cibles au niveau de l'enveloppe nucléaire.
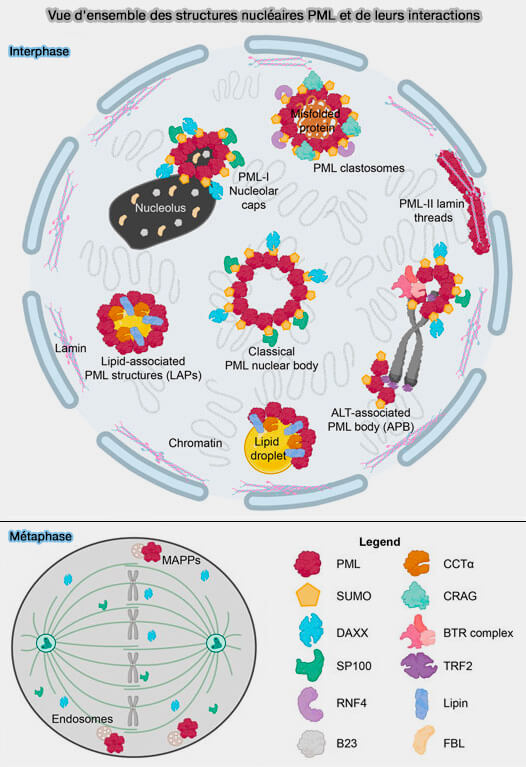
Vue d'ensemble des structures nucléaires PML et de leurs interactions
(Figure : vetopsy.fr d'après McPhee et coll)
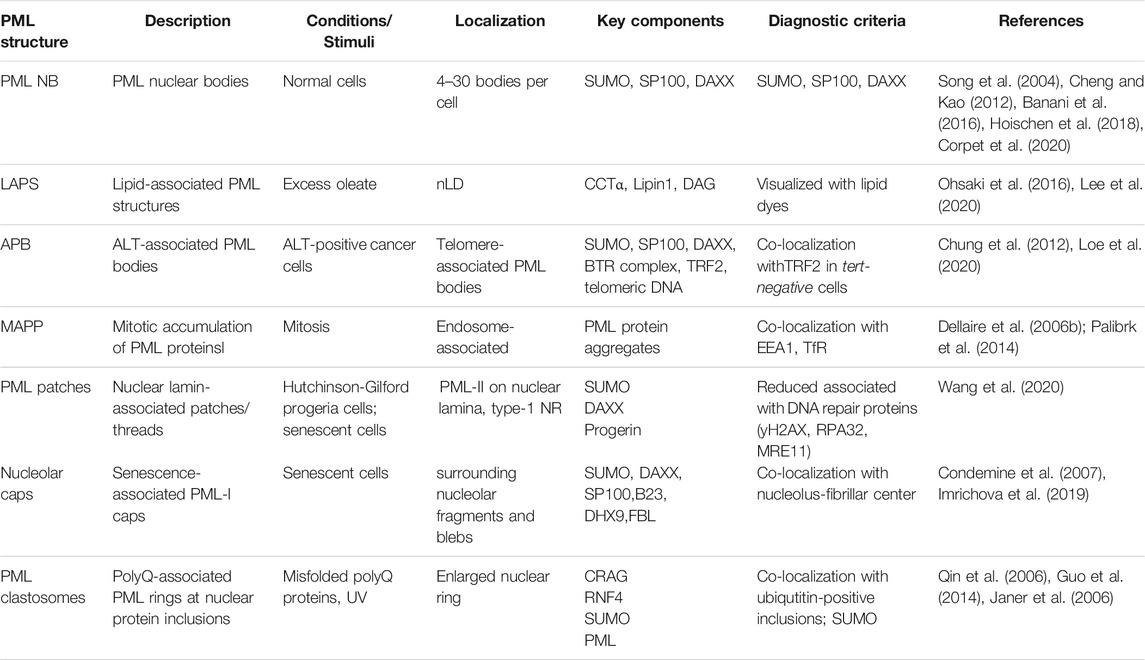
3. Dans cet excellent article traite de l'association des nLD avec la PML (ProMyelocytic Leukemia protein) pour former des LAPS (Lipid-Associated PML Structures) qui sont distincts des corps nucléaires canoniques des PML (NB) car ils manquent de protéines clés et de modifications associées à ces NB ( sous-structures contenant de la PML)
La PML est un régulateur clé des événements de signalisation nucléaire et les PML NB sont des sites de régulation génique et de modification post-traductionnelle des facteurs de transcription.
La sous-fraction des nLD qui forment les LAPS pourrait réguler les réponses au stress lipidique grâce à leur recrutement et à leur rétention de la protéine PML.
Les nLD et les LAPS possèdent des enzymes biosynthétiques lipidiques à leur surface, ce qui suggère qu'ils sont des sites actifs pour la synthèse des phospholipides nucléaires et des triacylglycérols ainsi que pour la régulation globale des lipides.






 sous-structures contenant de la PML)
sous-structures contenant de la PML)