a. L'activation du CASM par les agonistes TRPML1 ou par d'autres médicaments lysosomotropes, active le TFEB indépendamment de la cible mécaniste de l'activité de la rapamycine (mTOR) et de la régulation des nutriments.
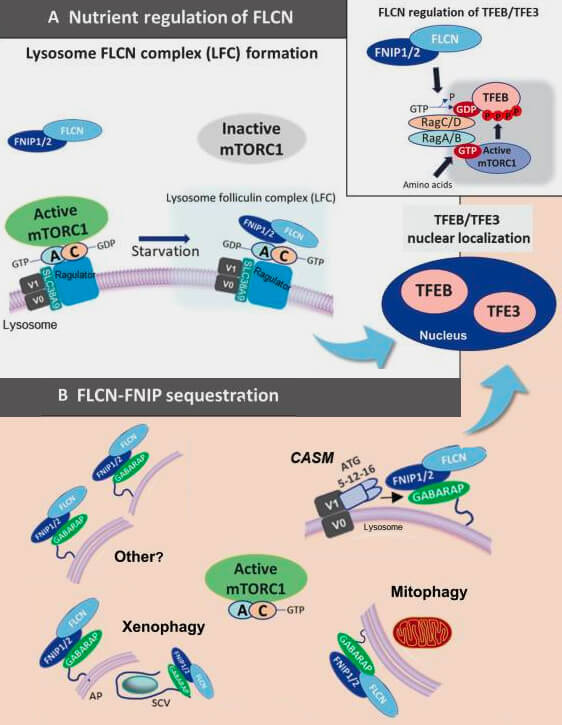
Séquestration de FLNC/FNIP par la CASM
(Figure : vetopsy.fr d'après Goodwin et coll)
b. Ce mécanisme alternatif repose sur GABARAP, et non sur LC3, qui se lie directement à un motif LIR au complexe folliculine (FLCN)/FNIP (folliculin-interacting protein).
La lipidation de GABARAP pendant la CASM séquestre ce complexe hors du cytosol, soulageant son effet inhibiteur sur les Rag guanosine triphosphatases (GTPases) et permettant ainsi l'activation du TFEB/TFE3 afin de contrôler la biogenèse lysosomale.
c. Ainsi, l’axe GABARAP-FLCN/FNIP-TFEB sert de capteur moléculaire qui coordonne l’homéostasie lysosomale avec les perturbations et le flux de cargo au sein du réseau autophagie-lysosomale.
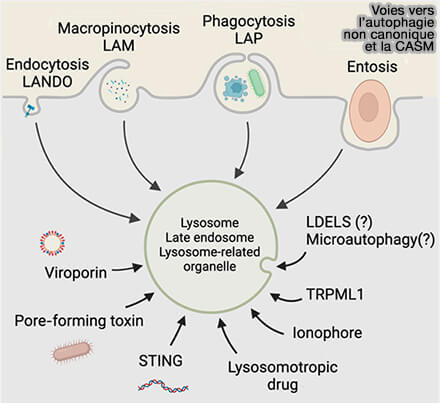
Agents pathogènes
Les agents pathogènes activent la CASM.
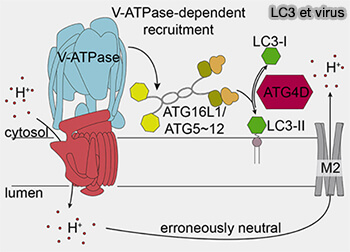
LC3 et virus
(Figure : vetopsy.fr d'après Ulfert et coll)
2. Les ionophores sont des espèces chimiques qui se lient de manière réversible aux ions.
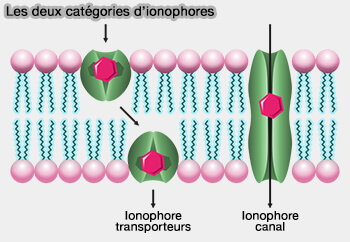
Les deux types d'ionophore
(Figure : vetopsy.fr d'après HovhannesKarapetyan)
a. De nombreux ionophores sont des entités liposolubles qui transportent des ions à travers la membrane cellulaire. Les ionophores catalysent le transport des ions à travers les membranes hydrophobes, comme les bicouches lipidiques présentes dans les cellules vivantes ou les vésicules synthétiques (liposomes).
b. Certains ionophores sont synthétisés par des micro-organismes pour importer des ions dans leurs cellules et comprennet deux catégories
Les ionophores transporteurs se lient à un ion particulier et protègent sa charge de l’environnement environnant, ce qui permet à l’ion de passer plus facilement à travers l’intérieur hydrophobe de la membrane lipidique,
Les ionophores porteurs sont des peptides comme la comme la valinomycine qui transporte un seul cation potassique, des protéines ou d’autres molécules.
Les ionophores à canaux transmembranaires forment un pore hydrophile dans la membrane, permettant aux ions de passer àsans entrer en contact avec l’intérieur hydrophobe de la membrane. [8]
Les ionophores formant des canaux sont généralement de grandes protéines.comme la gramicidine A et la nystatine.
Les PFT (Pore-Forming Toxins), i.e. protéines solubles, se lient aux récepteurs membranaires, ce qui conduit à leur oligomérisation et à l’insertion d’un pore aqueux dans la membrane plasmique (Role of Pore-Forming Toxins in Bacterial Infectious Diseases 2013).
Elles perturbent et endommagent les endomembranes, ce qui active alors la lysophagie, mais ont été aussi associées à une augmentation de la lipidation de la LC3.
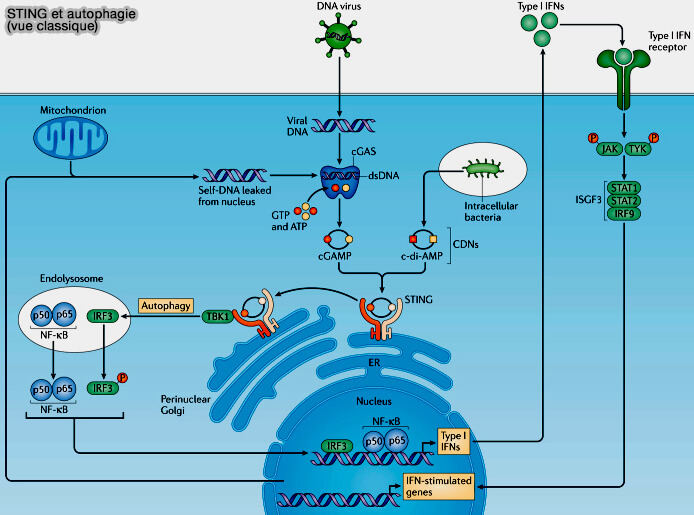
Voie cGAS-STING
1. La voie cGAS-STING (GMP-AMP cyclique synthase - STimulator of INterferon Genes) fait partie de la réponse immunitaire innée lors de la détection cytosolique d'un ADN à double brin dérivé :
1. L’engagement de la voie STING entraîne l’activation du facteur régulateur de l’interféron 3 (IRF3) pour la production d’interférons de type I et du facteur nucléaire κB (NFκB) et pour la production de cytokines inflammatoires.
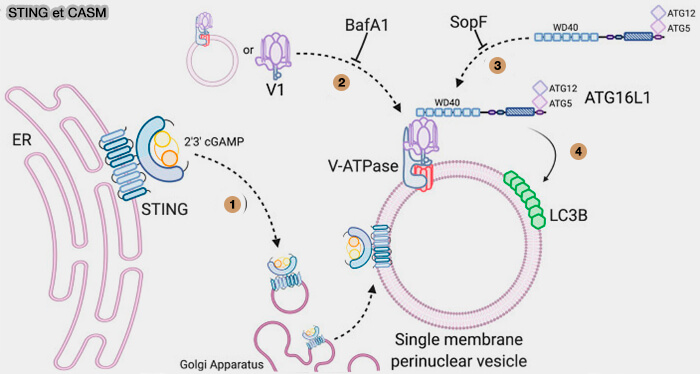
2. Le complexe V1 s’arrime aux domaines V0 résidents dans les vésicules périnucléaires, ou les vésicules avec des V-ATPases assemblées se redistribuent vers une formation plus dense dans la région périnucléaire.
4. LC3B est conjuguée à la phosphatidyléthanolamine (PE) sur les vésicules périnucléaires à membrane unique par le complexe ATG16L1-ATG5-ATG12.
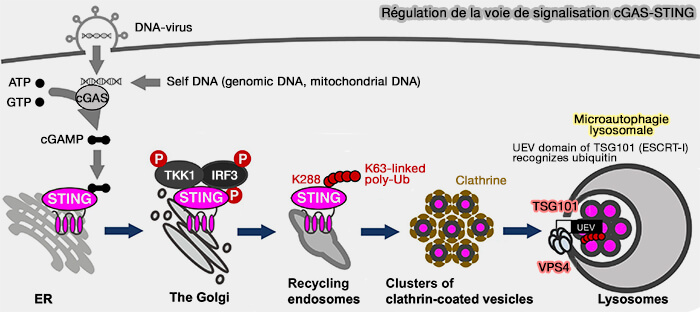
3. Toutefois, un autre processus, qui fait intervenir la microautophagie lysosomale, serait plus adapté à la dégradation de STING pour la prévention de l'activation immunitaire innée.
Régulation et dégradation de STING par microautophagie lysosomale
(Figure : vetopsy.fr d'après Kuchitssu et Tagushi)
a. Lors de la détection de l’ADN cytosolique par cGAS, STING se déplace :
Les KO de TSG101, sous-unité de ESCRT-I, ou Vps4 provoquent l'accumulation de vésicules dans le cytosol, conduisant à une réponse soutenue d'interféron de type I.




{kind=link}