


Autophagie
Autophagie non canonique
CASM : LDELS et conclusion générale
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L’autophagie et la production de vésicules extracellulaires (EV) sont interconnectées, et une partie des vésicules intraluminales (ILV) des endosomes ou corps multivésiculaires (MVE/MVB) peuvent suivre une voie appelée appelées LDELS (LC3-Dependent EV Loading and Secretion).
Sécrétion : LDELS (LC3-dependent extracellular vesicle
loading and secretion)
1. L’autophagie et la production de vésicules extracellulaires (EV) sont interconnectées.
a. Un complexe entre ATG12-ATG3 interagit avec la protéine accessoire ALIX du complexe ESCRT pour réguler une petite libération d'EV (ATG12-ATG3 Interacts with Alix to Promote Basal Autophagic Flux and Late Endosome Function 2015).
(Figure : vetopsy.fr d'après Guo et coll)
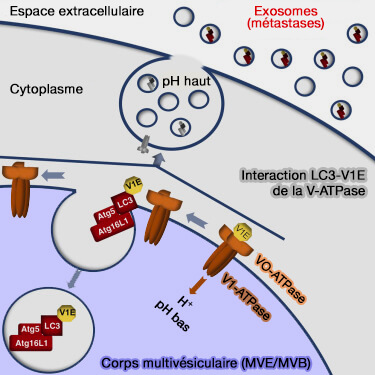
b. ATG5 et LC3 régulent de manière coordonnée l'acidification des endosomes tardifs ou corps multivésiculaires (MVE/MVB) et la production d'exosomes (Atg5 Disassociates the V1V0-ATPase to Promote Exosome Production and Tumor Metastasis Independent of Canonical Macroautophagy 2017).
- La production d’exosomes est fortement réduite dans les cellules dépourvues d’ATG5 et d’ATG16L1, mais indépendamment d’ATG7 et de l’autophagie canonique.
- ATG5 diminue spécifiquement l’acidification des endosomes tardifs où les exosomes sont produits, perturbant le complexe V-ATPase en éliminant un composant régulateur, ATP6V1E1, dans les exosomes.
c. En outre, les endosomes tardifs peuvent fusionner à des autophagosomes immatures.
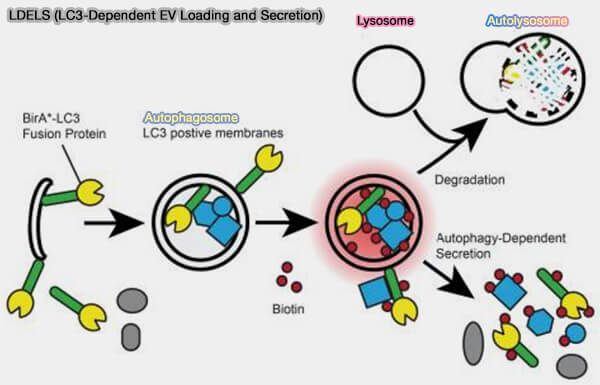
2. Une partie des vésicules sécrétées comme les vésicules intraluminales (ILV) des endosomes ou corps multivésiculaires (MVE/MVB) suit une voie appelée LDELS ou LC3-Dependent EV Loading and Secretion (The LC3-Conjugation Machinery Specifies the Loading of RNA-Binding Proteins into Extracellular Vesicles 2020).
(Figure : vetopsy.fr d'après Leidal et coll)
a. LC3/ATG8 et la machine de conversion LC3 capturent directement les RBP (RNA-Binding Protein), qui sont les cargos les plus répandus dans le LDELS et qui contiennent des motifs LIR.
- Elle les intègre dans des ILV grâce à un pool de LC3-II situé sur la membrane des MVB.
- Les ILV sont ensuite libérées sous forme d'EV via la fusion MVB avec la membrane plasmique.
b. La LDELS nécessite également le recrutement de FAN dépendant de nSMase2 et de LC3.
Les céramides produits par la nSMase2 peuvent entraîner la formation de microdomaines membranaires qui imposent une courbure négative et facilitent le bourgeonnement intraluminal indépendant du complexe ESCRT au niveau de la membrane des MVB (Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes 2008).
(Figure : vetopsy.fr d'après Lee et coll)
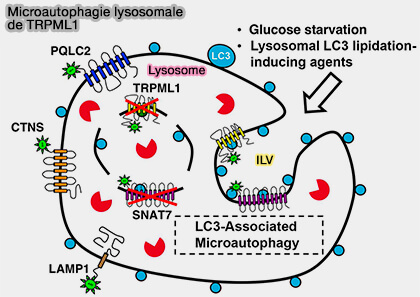
Remarque : la membrane lysosomale et plusieurs protéines membranaires lysosomales, comme TRPML1, peuvent être internalisées et dégradées dans la lumière lysosomale ( microautophagie lysosomale).
microautophagie lysosomale).
Conclusion générale sur la CASM
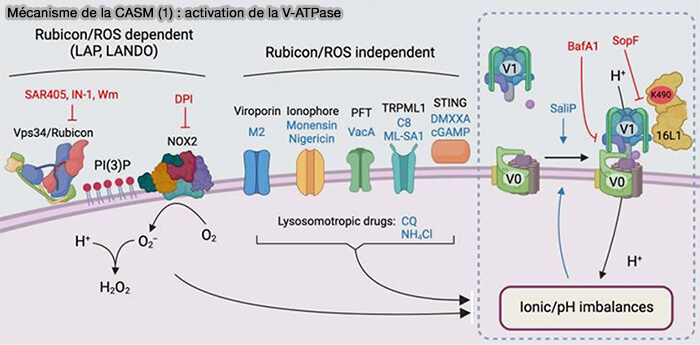
1. Différents stimuli convergent pour modifier l’équilibre ionique et le pH de la vésicule endolysosomale cible.
- La phagocytose associée à LC3 (LAP) et l'endocytose associée à LC3 (LANDO) dépendent de l’activation de NOX2, une NADPH oxydase, médiée par Vps34/Rubicon pour induire la production de ROS, qui consomment des protons et augmentent le pH.
- De multiples stimuli, y compris les viroporines, les ionophores, les PFT (Pore-Forming Toxins), les médicaments lysosomotropes, les agonistes de TRPML1 et les agonistes STING, entraînent tous des altérations des équilibres ioniques luminaux et du pH indépendamment de Rubicon et des ROS.
(Figure : vetopsy.fr d'après Durgan et Florey)
2. Les perturbations du pH endolysosomal sont détectées par des mécanismes inconnus et conduisent à une association accrue des sous-unités V0–V1 de la V-ATPase.
3. L’augmentation de l’association V0-V1 entraîne VAIL (V-ATPase-ATG16L1-induced LC3 lipidation) et le recrutement du complexe ATG16L1 via son domaine C-terminal WD40.
Les systèmes de conjugaison de type ubiquitine liés à l’autophagie ciblent ensuite les protéines ATG8 pour la lipidation en phosphatidyléthanolamine (PE) ou en phosphatidylsérine (PS) sur la membrane endolysosomale.
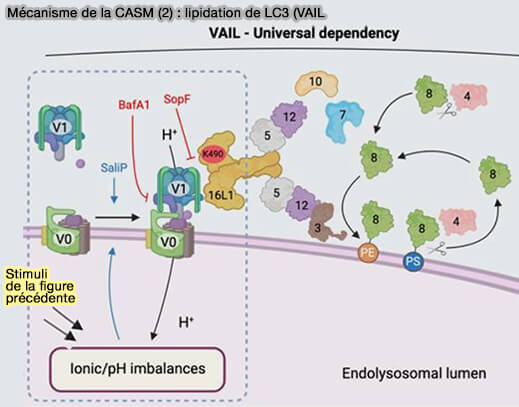
(Figure : vetopsy.fr d'après Durgan et Florey)
4. Les caractéristiques suivantes pourraient être considérées comme diagnostiques pour la CASM.
- 1. Conjugaison de l'ATG8 à une membrane simple (CASM) plutôt qu'à un autophagosome à double membrane.
- 2. Dépendance à l'égard des machines de conjugaison ATG8 de base (ATG3/4/5/7/10/12/16L1), associée à l'indépendance vis-à-vis des régulateurs de macro-autophagie en amont (ULK2, FIP200, ATG13/, ATG9, ATG14 et WIPI2).
- 3. Dépendance vis-à-vis du domaine WD40 d'ATG16L1 et du résidu K499, associée à l'indépendance de son domaine FBD de liaison à FIP200.
- 4. Induction d'une conjugaison alternative de l'ATG8 à PS plutôt qu'exclusivement à PE.
- 5. Dépendance à l'axe V-ATPase-ATG16L1, comme en témoigne l'inhibition par la bafilomycine A1 (BafA1) et/ou SopF.
- 6. Si la dépendance à Rubicon et NOX2 peut être utilisée pour impliquer LAP (et LANDO), cela ne doit pas être utilisé pour exclure d'autres formes d'autophagie non canonique et de CASM qui se couplent à l'axe V-ATPase-ATG16L1 indépendamment.
Retour à l'autophagie
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation