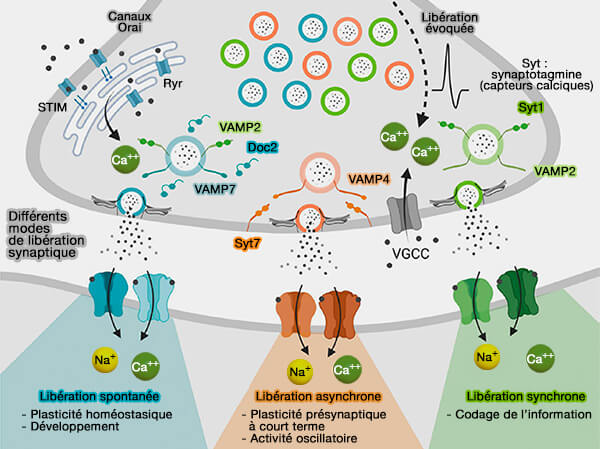
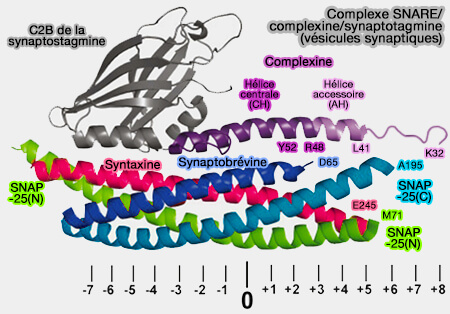
Les complexines et les synaptotagmines régulateurs ne sont pas nécessaires à la fusion en soi, mais ils permettent l'accumulation, au niveau de la membrane plasmique, d'une réserve (pool) de vésicules prêtes à fusionner en attente du signal, i.e. entrée de calcium pour une libération rapide et synchrone.
Vue d'ensemble des rôles de la complexine
Les rôles de la complexine sont difficiles à analyser, car les modèles ont fortement évolué au cours du temps et sont dépendants du mode de transmission des neurotransmetteurs et de ses interactions avec les synaptotagmines. Cplx a plusieurs rôles :
La principale différence entre Cplx3 et d'autres isoformes de mammifères se trouve dans l'extrémité C-terminale, ce qui suggère que cette région abrite des déterminants critiques pour le verrouillage de la fusion.
Cette composante plus lente de la libération se produit lorsque les niveaux de Ca++ chutent par rapport à leur concentration maximale, ce qui entraîne la fusion synchrone.
Le verrouillage de Cplx agit de manière optimale à une concentration de Ca++ plus élevée, i.e. il agit en partie en régulant la sensibilité calcique de la libération des SV.
l'hélice accessoire (AH) se projette à un angle de 45° pour lier un complexe SNARE partiel voisin.
Modèle zigzag de la complexine
(Figure : vetopsy.fr d'après Kümmel et coll)
1. Ce mode permettrait à Cplx de relier des complexes SNARE partiels dans une chaîne en zigzag alternée, prise en sandwich entre les vésicules synaptiques (VS) ancrées et la membrane présynaptique pour verrouiller la libération et inhiber la fusion avant l'influx de Ca++.
La palmitoylation de SNAP-25 limite la distance entre le zig-zag et la membrane plasmique.
La distance entre le plan en zigzag et la vésicule synaptique doit être inférieure à 110 Å, la distance maximale parcourue par le lien v-SNARE.
AH se lie probablement à l'extrémité C-terminale de VAMP2, déstabilisant ainsi l'étape finale de la fermeture éclair SNARE en excluant VAMP2 de l'extrémité C-terminale du complexe SNARE.
Modèle de compétition modifié
(Figure : vetopsy.fr d'après Malsam et coll)
AH crée une barrière entre le faisceau SNARE et la bicouche de la vésicule synaptique (VS).
L'attachement de AH à une VS favoriserait la séparation de AH du faisceau SNARE et, respectivement, stabiliserait l'état partiellement dézippé de VAMP2.
b. Le verrouillage de la fusion peut être amélioré en renforçant les interactions de AH avec la bicouche VS en remplaçant les résidus Cplx qui font face au faisceau SNARE par des résidus plus hydrophobes, tels que Ala, Phe, Trp ou Leu. De plus, le modèle prédit également que la pince de fusion peut être renforcée en affaiblissant les interactions entre l'AH de Cplx et le complexe SNARE.
La synaptotagmine-1 (Syt1) et la complexine coopèrent pour empêcher la fermeture éclair complète des SNARE pendant l'amorçage des vésicules synaptiques et activent plus tard la fusion complète après l'influx de Ca++.
Facilitation de la fusion comme fonction secondaire ?
La grande majorité des systèmes modèles ainsi que les analyses in vitro indiquent une facilitation de la fusion par la complexine, qui complète le verrouillage de la libération spontanée ou qui pourrait même représenter sa fonction principale en fonction du modèle ( voir revue).
Modèle de facilitation de la fusion par la complexine
(Figure : vetopsy.fr d'après Xue et coll)
1. L'extrémité N-terminale de la complexine, chez les vertébrés, faciliterait la fusion.
Cette action facilitatrice semble mécaniquement indépendante et même séparable de la fonction de verrouillage de la complexine, qui est supposément médiée par l'hélice α accessoire avec l'extrémité C-terminale.
Cependant, chez les invertébrés, le rôle mécaniste des domaines de complexation peut s'écarter de ce modèle.
Extrémité N-terminale de la complexine
(Figure : vetopsy.fr d'après Xue et coll)
Le motif N-terminal impliqué formerait un segment alpha-hélicoïdal amphipathique qui se lie à l'extrémité C-terminale du complexe SNARE.
L'effet facilitateur observé sur la libération évoquée peut s'expliquer par la liaison de ce motif hélicoïdal au complexe SNARE qui pourrait fournir un support conformationnel à l'assemblage de l'extrémité C-terminale au cours des dernières étapes de l'exocytose.




 modèles de régulation des SNARE par la complexine/synaptotagmine).
modèles de régulation des SNARE par la complexine/synaptotagmine).