un domaine transmembranaire N-terminal (1-58), qui lie Syt à la vésicule synaptique,
un lien (linker) juxtamembranaire (80-139),
Domaines C2 de Syt1
(Figure : vetopsy.fr d'après Xue et coll)deux domaines C2 cytoplasmiques C-terminaux en tandem, appelés C2A (140-254) et C2B,(272-479), homologues à ceux de la protéine kinase C (PKC), séparés par un domaine de liaison flexible à 9 résidus.
Ces domaines interagissent avec la membrane et la pénètrent.
C2B est le domaine fonctionnel principal de la synaptotagmine.
La mutation d'un résidu de la pointe hydrophobe de la boucle 3 de C2B, i.e. l'isoleucine (I420) de Syt1, qui pénètre dans les membranes chargées négativement, est mortelle pour l'embryon et provoque une diminution de la libération évoquée plus sévère que celle observée chez les mutants nuls de Syt1.
Résidus hydrophobes ancrés dans la membrane
(Figure : vetopsy.fr)
La liaison Ca++ neutralise efficacement la répulsion électrostatique entre la membrane cible et la poche de liaison calcique, permettant aux résidus non polaires situés à l'extrémité de chaque boucle de calcium de s'insérer dans la membrane (Ca2+-Triggered Simultaneous Membrane Penetration of the Tandem C2-Domains of Synaptotagmin I 2006).
4. Deux résidus hydrophobes hautement conservés sont localisés aux extrémités de chaque poche :
un dans la boucle 1, adjacent au premier résidu d'aspartate, M173 dans C2A et V304 dans C2B,
Ces résidus sont partiellement insérés dans le feuillet interne de la membrane plasmique (10 Å de profondeur) et peuvent induire une déformation locale de la membrane plasmique (How Synaptotagmin Promotes Membrane Fusion 2007).
Différences entre C2A et C2B
Hélice HB de C2B de Syt1
(Figure : vetopsy.fr d'après Wu et Schulten)
Hélices de C2B
1. La région C-terminale de C2B, contrairement à C2A, contient deux hélices.
b. Lors d'expériences sur les granules sécrétoires purifiés, l'élimination de PIP2 réduit leur liaison à la membrane et l'efficacité de la fusion, ralentit le moment de la fusion ou le mode de libération, modifie la conformation SNARE et réduit la cinétique de fusion.
L'élimination de PIP2 produit également l'ouverture du pore de fusion similaire à celui des mutants RQ et RQRQ.
3. En présence de Ca++, les boucles de liaison au Ca++ et la face polybasique fonctionnent en coopération pour conduire l'association membranaire du domaine C2B.
Remarque : la mutation RQ déplace l'apex vers la phase aqueuse de ~5 Å, la double mutation RQRQ élimine l'interaction membranaire.
Le domaine C2B établit un contact membranaire simultané au niveau de l'apex d'arginine, de la face polybasique et des boucles de liaison au Ca++, ce qui ne serait possible qu'avec une surface courbe (Computational Lipidomics of the Neuronal Plasma Membrane 2017).
La liaison membranaire des domaines C2 est nécessaire pour initier la fusion, mais les interactions membranaires créées par l'apex d'arginine sont nécessaires pour en déterminer le bon moment et l'ouverture rapide du pore de fusion.
Interactions apex arginine/SNARE
(Figure : vetopsy.fr d'après Nyenhuis et coll)
2. L'apex s'associe également aux SNARE par T285, F349 et E350, ce qui est un peu différent du modèle tripartite ( région II de l'interface primaire).
Remarque 1 : la mutation RQ déplace l'apex vers la phase aqueuse de ~5 Å, la double mutation RQRQ élimine l'interaction membranaire.
Syt7 démontre un mode de libération qui est significativement différent de Syt1, i.e. le moment de l'événement de fusion semble être ralenti.
Toutefois, la mutation simultanée des sites de la boucle 1 et de la boucle 3 de C2A a entraîné une abolition presque complète de la libération évoquée, i.e. plus sévère que celle observée chez les mutants nuls de la Syt.
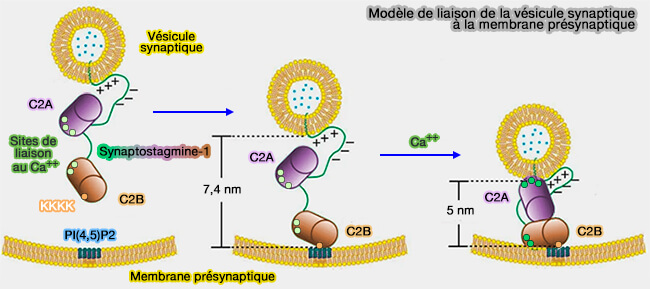
Modèle de liaison de la vésicule synaptique à la membrane présynaptique
(Figure : vetopsy.fr d'après Lin et coll)
Le domaine C2A serait un composant essentiel pour traduire la fonction de commutation électrostatique de la synaptotagmine en fusion de la vésicule avec la membrane présynaptique. Les deux domaines coopèrent pour positionner Syt1 dans la configuration la plus favorable pour une énergie de liaison maximisée.
L'attachement transmembranaire aux vésicules synaptiques par le maintien de la longueur du linker juxtamembranaire et l'orientation du domaine C2 sont essentiels pour que Syt1 régule la neurotransmission.
Cette attache vésiculaire permet aux domaines C2A-C2B de Syt1 de réguler plusieurs modes de libération de neurotransmetteurs, i.e. libération synchrone ou libération asynchrone.
Lien (linker) de Syt1
(Figure : vetopsy.fr d'après Lai et coll)











 interface tripartite).
interface tripartite).