Les V-ATPases, complexes multimériques qui fonctionnent comme un nanomoteur rotatif de pompage de protons, sont composées de deux sous-unités.
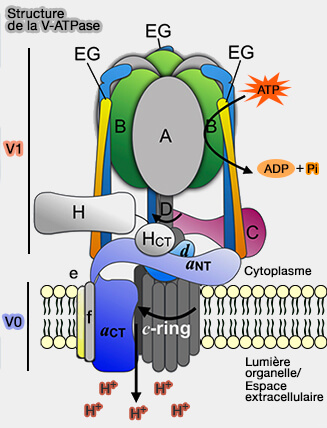
Structure des V-ATPases
1. Le secteur transmembranaire (V0), formé de six sous-unités différentes (ac4c′c″de), est responsable de la translocation des protons.Le chiffre en indice indique le nombre d'exemplaires de la sous-unité.
Structure de la V-ATPase
(Figure : vetopsy.fr d'après Yao et coll)
a. Les sous-unités protéolipidiques (c, c′ et c″) sont organisées en un anneau contenant des copies uniques des sous-unités c′ et c″ et des copies multiples de la sous-unité c.
b. TM1 de la sous-unité c″ semble être dispensable pour la translocation.
La sous-unité a de V0 fournit des canaux d'accès (hémi-canaux) qui permettent aux protons d'atteindre et de quitter ces résidus acides enfouis sur le cycle protéolipidique.
2. Le secteur V1 cytoplasmique, constitué de huit sous-unités différentes avec une stoechiométrie définie (A3B3CDEFG2H1-2), est responsable de l'hydrolyse de l'ATP.
a. L'hydrolyse de l'ATP se produit au niveau de sites catalytiques situés à l'interface des sous-unités A et B, qui sont chacune présentes en trois exemplaires par complexe et disposées en alternance dans un anneau. La plupart des résidus du site catalytique sont apportés par la sous-unité A.
b. Un deuxième ensemble de sites de liaison de nucléotides est situé à l'autre interface de sous-unité A/B (appelés sites " non catalytiques "), qui sont composés principalement de résidus de sous-unité B et peuvent fonctionner pour réguler l'activité.
Dans cette région, un insert de 90 acides aminés " dépasse " de la sous-unité A dans la tête hexamérique A3B3 et pourrait contrôler l'assemblage dépendant du glucose du complexe.
3. L'hexamère catalytique A3B3 est relié à V0 par des tiges.
a. La tige centrale, formée par les sous-unités D et F de V0 et d de V1, fonctionne comme un " rotor ".
Elle s'étend de l'anneau protéolipidique jusqu'au centre de la tête hexamérique A3B3 et est attachée à un anneau de sous-unités hydrophobes (c, c,′ c″)
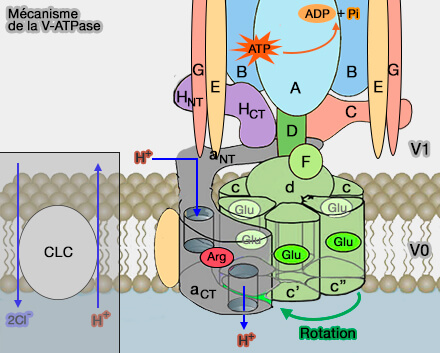
Mécanisme de la V-ATPase
(Figure : vetopsy.fr d'après plusieurs sources)
b. Les deux tiges périphériques forment un " stator ", i.e. partie fixe par opposition au rotor, partie mobile.
Ces tiges sont formées par le domaine N-terminal cytosolique de la sous-unité a avec les sous-unités C, E, G et H attachées à l'hexamère A3B3 .
L'hydrolyse de l'ATP et le pompage de protons sont couplés par un mécanisme rotatif, c'est-à-dire la rotation d'un " rotor " par rapport à un " stator " dans la nano-machine.
Mécanisme des V-ATPases
1. L'hydrolyse de l'ATP au niveau des sites catalytiques de V1 entraîne une rotation dans le sens des aiguilles d'une montre de la tige centrale et de l'anneau des sous-unités protéolipidiques par rapport à la sous-unité a, " clouée " par les tiges périphériques qui empêchent sa rotation par rapport à A3B3.
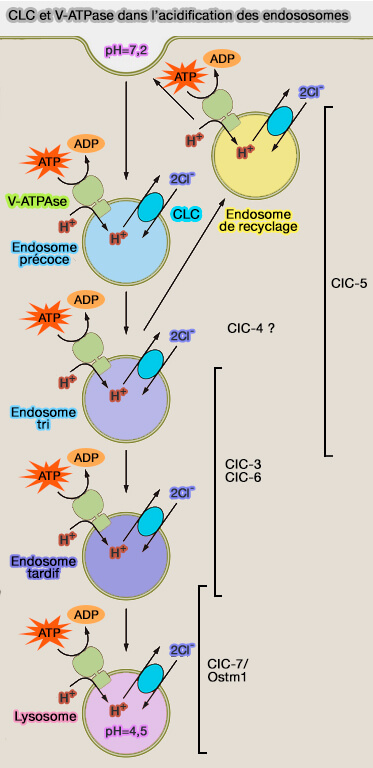
Un certain nombre de mécanismes sont utilisés pour réguler l'activité de la V-ATPase pour réguler le pH à la fois dans l'espace et dans le temps.
On peut prendre comme exemples de gradient de pH :
la maturation des endosomes dont le pH s'acidifie de l'endosome précoce aux lysosomes pour leur permettre d'assurer leurs fonctions,
le parcours des vésicules golgiennes, i.e. dissociation des enzymes lysosomales du récepteur du mannose-6-phosphate et le recyclage de ce récepteur vers le trans-Golgi.
Ces mécanismes, loin d'être tous décryptés, la complexité structurelle ainsi que les différents isoformes qui localisent la V-ATPase dans des environnements cellulaires ou des systèmes d'organes distincts, permettent une régulation différenciée.
Dissociation réversible des composants V1 et V0
La dissociation réversible des composants V1 et V0 est le mécanisme le plus important de régulation de l'activité de la V-ATPase, mécanisme qui annihile et l'hydrolyse de l'ATP et la translocation passive des protons qui sinon auraient des effet délétères.
Dissociation réversible de la V-ATPase
(Figure : vetopsy.fr d'après Yao et coll)
RAVE est composé de trois protéines (Rav1p, Rav2p et Skp1, un composant de l'ubiquitine ligase et semble se lier aux sous-unités E et G de V1 ainsi qu'à la sous-unité C (une sous-unité V1 qui se dissocie de V1 et V0 lors du désassemblage).
RAVE semble stabiliser le complexe V1 dissocié sous une forme compétente pour le réassemblage et assure la médiation de l'assemblage.
L'activité de la V-ATPase est en équilibre entre un état entièrement couplé et entièrement découplé.
Toutefois, le transport de protons peut être modifié en augmentant ou en diminuant l'efficacité de couplage par rapport à cet état de départ.
3. La formation de ponts disulfure réversibles au niveau du site catalytique de la sous-unité A permet une autre forme de régulation.
Dans la levure, la Cys-261 conservée du site catalytique et la Cys-539 dans la même sous-unité forment une liaison disulfure qui se traduit par inactivation réversible de la V-ATPase en bloquant l'hydrolyse de l'ATP (Site-directed Mutagenesis of the Yeast V-ATPase A Subunit 1997).
La V-ATPase est électrogénique, i.e. un afflux continu de protons à travers les membranes entraînerait une accumulation de charge positive toujours croissante dans la lumière des endosomes et des lysosomes.
2. Chez les mammifères, les protéines CLC forment une grande famille de neuf membres.
CLC3 à CLC7 se localisent sur les membranes du système endosomal-lysosomal.
2. Les CLC vésiculaires stimulent l'activité de la V-ATPase dans les compartiments endosomal-lysosomal, facilitant ainsi l'acidification.
Les CLC permettent l'influx de H+ contre un gradient de concentration, mais compensent en minimisant le gradient électrochimique, entraînant une régulation nette du pH des compartiments vésiculaires.
En retour, les courants ioniques CLC sont régulés par l'état de l'environnement intraluminal : par exemple, un faible pH luminal est capable d'ouvrir le déclenchement du canal CLC et les courants Cl−.