de tube, de pont et de tunnelmultimériques ou monomériques, ce qui a l'énorme avantage de déplacer le lipide dit par glissement (sliding en anglais) sans ou avec un minimum de mouvements des protéines, i.e. ce qui permet une augmentation substantielle de la vitesse du transfert.
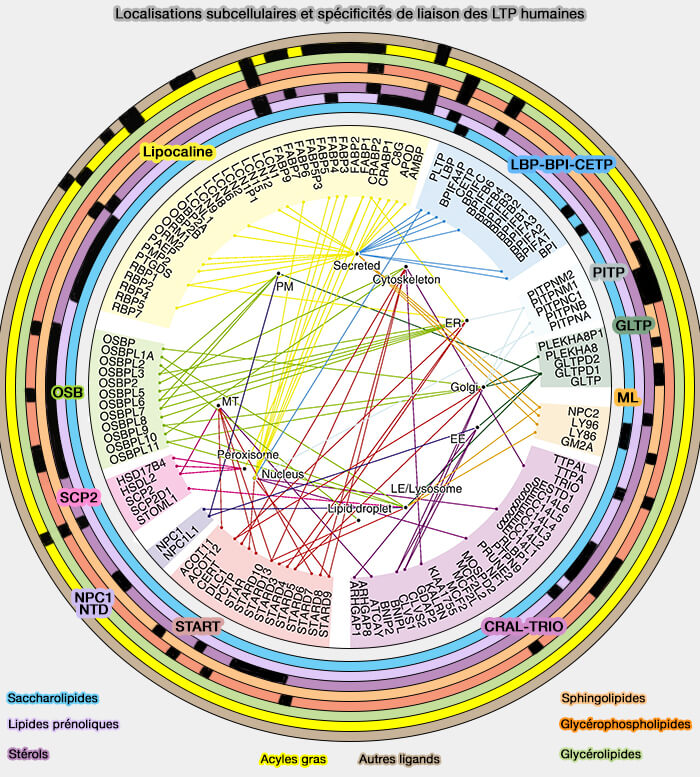
Localisations subcellulaires et spécificités de liaison des LTP humaines
(Figure : vetopsy.fr d'après Chiapparino et coll)
Vue d'ensemble des transferts lipidiques par navette
1. La plupart des transporteurs connus fonctionnent comme des navettes qui transportent des molécules lipidiques uniques entre les membranes :
soit par un domaine simple,
soit par plusieurs domaines comprenant des domaines d'échange lipidique et/ou d'autres modules protéiques impliqués dans la liaison membranaire spécifique et/ou l'interaction avec d'autres protéines associées ou intégrées à la membrane, déterminant ainsi leur emplacement cellulaire et leurs capacités de connection.
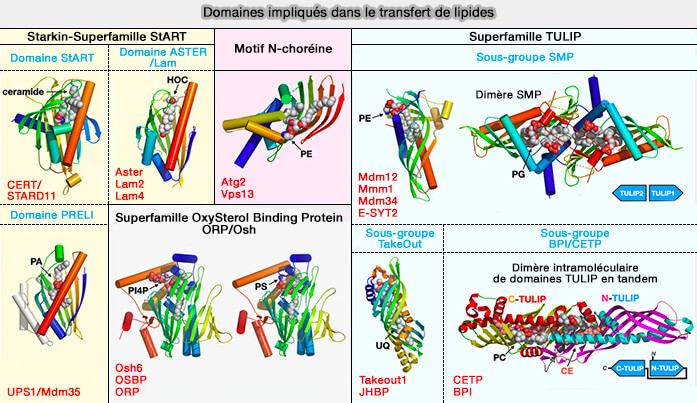
Domaines impliqués dans le transfert des lipides
(Figure : vetopsy.fr d'après Egea)
2. Leurs domaines de transfert lipidique ont généralement une taille d'environ 150 à 300 acides aminés.
a. Ils sont formés de feuillets β courbés pour se plier en structures en forme de panier, de tonneau ou de coupe et former des cavités hydrophobes qui protègent les lipides de l'environnement pendant le transport, i.e. leur énergie est bien inférieure à celle des milieux aqueux ( coût énergétique du transfert).
La sélection du lipide s'effectue par son groupe de tête et sa chaîne acyle (R-C(=O)-) grâce à une interaction spécifique avec des résidus distincts dans la cavité.
Toutefois, certains LTP peuvent se lier à deux lipides à groupes de tête différents pour effectuer des échanges.
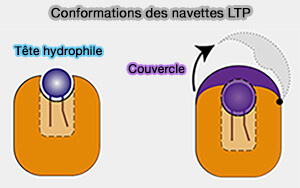
c. Certaines de ces structures ressemblent à des paniers, scellés à une extrémité et dotés de boucles flexibles ou de petites hélices agissant comme des couvercles ou des portes à leur entrée pour piéger et libérer de manière réversible les ligands lipidiques et fonctionner comme des navettes.
Conformation des navettes LTP
(Figure : vetopsy.fr d'après Wong et coll)
a. La cavité hydrophobe interne renferme une partie ou la totalité du ligand lipidique pour le protéger du milieu aqueux.
b. Ces LTP peuvent :
soit exposer le groupe de tête hydrophile,
soit posséder un couvercle (lid en anglais) qui peut s'ouvrir et se fermer par changement conformationnel, i.e. le groupe de tête hydrophile peut être soit proximal, soit distal.
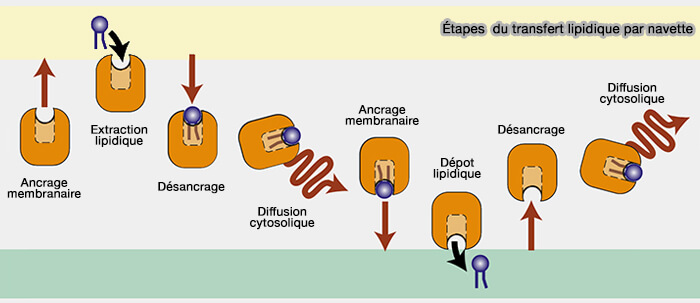
4. Ce transfert lipidique nécessite 8 étapes.
a. Sur le compartiment donneur, il faut :
un ancrage membranaire,
une extraction lipidique,
un désancrage
une diffusion cytosolique.
b. Sur le compartiment receveur, les étapes sont semblables, excepté le dépôt au lieu de l'extraction.
Étapes du transfert lipidique par navette
(Figure : vetopsy.fr d'après Wong et coll)
c. La LTP, après le transfert du lipide du donneur à l'accepteur, retourne :
soit vide pour réaliser le trafic lipidique,
soit avec un lipide différent lors d'échange, comme dans le cas de LTP bispécifique.
Nous ne pouvons pas développer l'étude de toutes les LTP : vous pouvez vous référer à plusieurs chapitres de vetopsy.fr et à la bibliographie dédiée.
La disposition particulière des éléments de structure secondaire des base d'hélices α et des feuillets β donne naissance à une structure tridimensionnelle constituée d'une cavité hydrophobe, ouverte sur l'extérieur.
Les protéines avec le pli Bet v1 se trouvent dans tous les règnes et se diversifient en de nombreuses familles avec une faible similarité de séquence, mais avec un pli commun au cours de l'évolution.
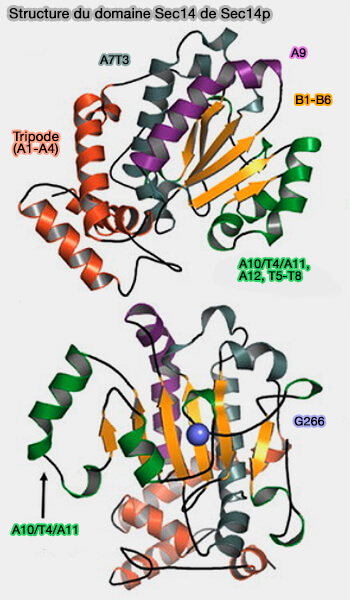
a. Ce domaine comprend 12 hélices α (A1-A12), 6 brins β (B1-B6) et 8 hélice-310 (T1-T8).
Le lobe N-terminal est constitué de quatre hélices α antiparallèles, A1-4 de 129 résidus, formant forme un tripode pour le ciblage de l'appareil de Golgi.
Le lobe C-terminal, appelé domaine CRAL-TRIO, i.e. appelé ainsi pour Cellular RetinALdehyde-binding protein (CRALBP) et TRIple functional domain protein guanine exchange factor, définit la poche de liaison des phospholipides (PL).
Le plancher de la cavité de liaison Sec14p se compose de cinq brins β parallèles comprenant un feuillet β pris en sandwich par deux longues hélices α d'un côté et deux courtes hélices β plus une hélice-310 de l'autre.
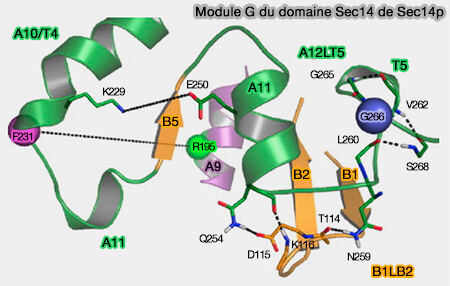
Module G du domaine Sec14 de Sec14p
(Figure : vetopsy.fr d'après Ryan et coll)une charnière qui lie les extrémités N- et C-terminales de la porte hélicoïdale, i.e. F212, Y213, K239, I240, et I242 (B4-B5) qui flanque A10/T4/A11,
un module de déclenchement, ou module G, i.e. B1LB2 and A12LT5, à travers lequel les informations conformationnelles sont transduites à la charnière.




 coût énergétique du transfert).
coût énergétique du transfert).