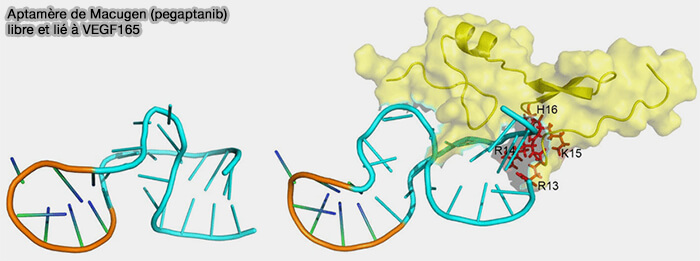
Acides nucléiques
ARN : types et fonctions
ARN non codants régulateurs de l’expression génique
2. Régulation indépendante de l’interférence par l’ARN
lncRNA, circRNA et riboswitch
Les lncRNA, circRNA et riboswitch sont des ARN régulateurs qui modulent l’expression génique par des mécanismes indépendants de l’interférence par l’ARN, impliquant des interactions structurales et fonctionnelles variées.
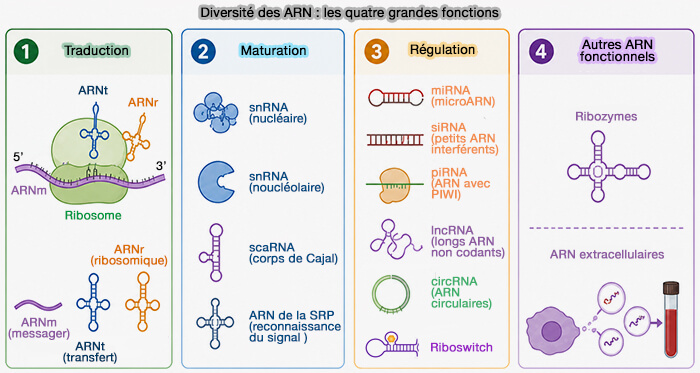
Les quatre grandes familles d'ARN
(Figure : vetopsy.fr)
1. Ils comprennent notamment :
les ARN longs non codants (lncRNA), qui interviennent dans la régulation transcriptionnelle, l’organisation de la chromatine, la structuration nucléaire ou le contrôle post-transcriptionnel,
les ARN circulaires (circRNA), ARN circulaires pouvant agir comme régulateurs, éponges à miRNA ou, plus rarement, comme matrices traduites,
les riboswitch, structures régulatrices présentes principalement dans certains ARN messagers bactériens, capables de modifier la conformation de l’ARN en réponse à la liaison directe d’un métabolite, contrôlant ainsi la transcription, la traduction ou la stabilité de l’ARN.
2. Ces petits ARN régulateurs agissent au sein de complexes effecteurs qui utilisent l’appariement de bases pour reconnaître des ARN cibles et provoquer leur dégradation ou leur inhibition traductionnelle.
Malgré ces similitudes structurales, ces ARN ne sont pas traduits et exercent essentiellement des fonctions de régulation de l’expression génique.
Les lncRNA participent à la régulation de l’expression génique par plusieurs mécanismes moléculaires reposant sur leur capacité à interagir avec l’ADN, les ARN et de nombreuses protéines régulatrices.
Remarque : certains ARN nucléaires, désignés comme Chromatin-Associated RNAs (caRNA), restent étroitement associés à la chromatine et participent à son organisation tridimensionnelle et au recrutement de complexes régulateurs impliqués dans la régulation de l’expression génique
Ils correspondent :
le plus souvent à des lncRNA, comme Xist, HOTAIR ou NEAT1,
1. L’un des principaux modes d’action des lncRNA repose sur leur capacité à servir de plateformes moléculaires permettant l’assemblage de complexes ribonucléoprotéiques régulateurs.
a. La structure secondaire et tertiaire de ces ARN crée des surfaces d’interaction capables de lier simultanément plusieurs protéines régulatrices.
Cette propriété permet aux lncRNA d’agir comme molécules d’échafaudage qui rapprochent des enzymes de modification de la chromatine, des facteurs de transcription ou des complexes de remodelage nucléosomique, facilitant ainsi leur action coordonnée sur des régions génomiques spécifiques.
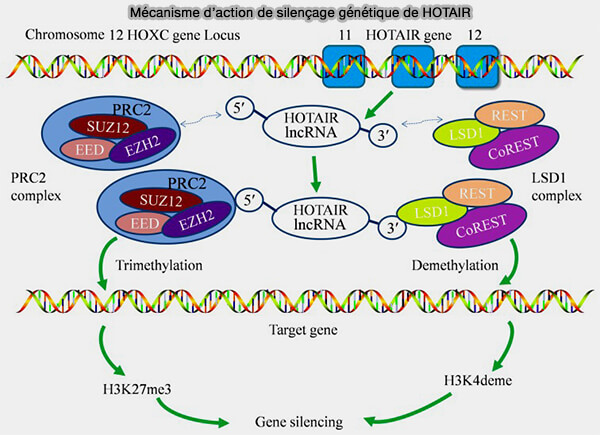
Mécanisme d'action du silençage génétique par la lncRNA Hotair
(Figure : vetopsy.fr d'après Wu et coll)
a. L’appariement direct avec des séquences d’ADN ou l’interaction avec des protéines associées à la chromatine permet de positionner les complexes enzymatiques sur des promoteurs ou de domaines nucléaires particuliers.
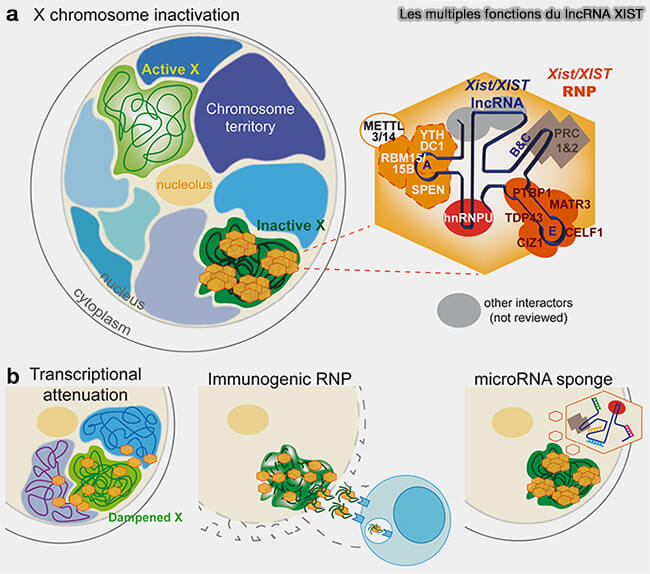
Les multiples fonction du lncRNA Xist
(Figure : vetopsy.fr d'après Morey et coll)
b. Le lncRNA Xist (X-inactive specific transcript) est l’un des exemples les mieux caractérisés de lncRNA régulateur de la chromatine, composée de plusieurs régions répétées capables de recruter des partenaires protéiques distincts, montrant ainsi comment un même ARN peut coordonner différentes étapes du silencing chromatinien (Unleashing XIST from X-chromosome inactivation 2025).
Ce long ARN est transcrit à partir du centre d’inactivation du chromosome X (X-inactivation center, XIC) et reste et reste majoritairement associé au chromosome dont il est issu.
3. Les lncRNA peuvent également intervenir dans l’organisation tridimensionnelle du génome.
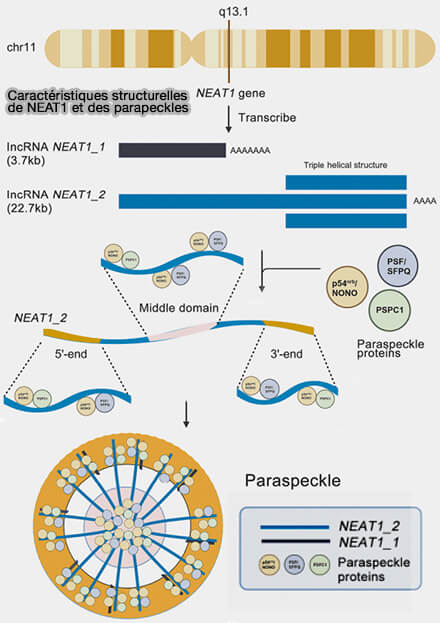
Caractéristiques structurelles de NEAT1 et des paraspeckles
(Figure : vetopsy.fr d'après Cao et coll)
En interagissant simultanément avec plusieurs régions chromosomiques ou avec des protéines structurales de la chromatine, ces ARN contribuent à la formation de domaines nucléaires fonctionnels et à la stabilisation d’interactions entre régions génomiques distantes, favorisant ainsi la régulation coordonnée de groupes de gènes au sein du noyau.
4. Enfin, certains lncRNA exercent leurs fonctions au niveau post-transcriptionnel en interagissant directement avec d’autres ARN.
a. Ces interactions peuvent modifier la stabilité, la localisation ou la traduction des ARN messagers, notamment en modulant l’accès de protéines régulatrices ou de microARN à leurs cibles.
En contenant plusieurs sites de liaison pour un même microARN, ces lncRNA peuvent agir comme " éponges " moléculaires (sponge) et limiter l’interaction de ces microARN avec leurs ARN messagers cibles.
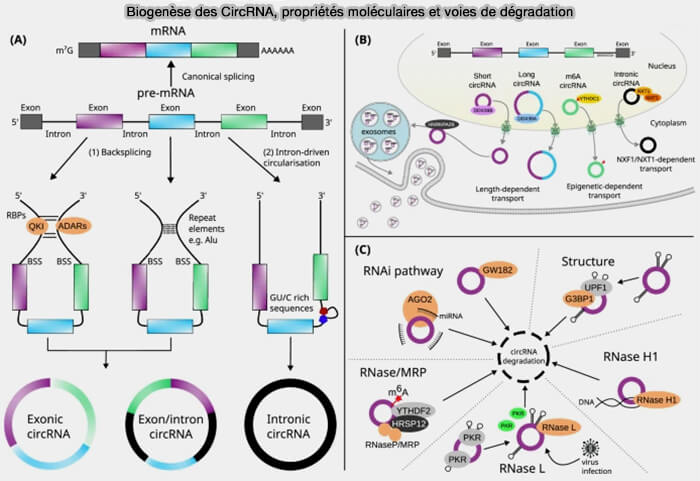
Les ARN circulaires (circRNA) constituent une classe particulière d’ARN non codants caractérisés par une structure covalente fermée dépourvue d’extrémités 5′ et 3′ libres.
1. Cette conformation résulte d’un mécanisme d’épissage non canonique appelé rétro-épissage (back-splicing), dans lequel un site donneur d’épissage en aval est relié à un site accepteur situé en amont, formant ainsi une molécule circulaire stablestable et une jonction caractéristique appelée jonction de rétro-épissage (back-splice junction).
La circularisation est souvent favorisée par des séquences répétées complémentaires présentes dans les introns flanquants, qui rapprochent les sites d’épissage et facilitent la formation de la boucle d’ARN. Chez l’humain, des éléments répétitifs de type ALU participent fréquemment à ce rapprochement des séquences impliquées dans le rétro-épissage (Circular RNAs are abundant, conserved, and associated with ALU repeats 2013).
b. La formation d’un circRNA résulte d’un rétro-épissage utilisant les mêmes sites d’épissage que ceux impliqués dans la production de l’ARNm linéaire.
Lorsqu’un exon est engagé dans un rétro-épissage pour former un circRNA, il n’est plus incorporé dans l’ARNm linéaire correspondant, ce qui peut modifier la proportion relative de transcrits linéaires et circulaires produits à partir du même gène.
Cette compétition entre épissage linéaire et circularisation constitue ainsi un niveau supplémentaire de régulation de l’expression du gène parental.
2. L’absence d’extrémités libres rend les circRNA particulièrement résistants aux exonucléases, ce qui leur confère une stabilité moléculaire élevée dans les cellules.
Cette stabilité explique leur accumulation dans certains tissus, notamment dans les neurones, où de nombreux circRNA présentent une expression spécifique du développement et du type cellulaire.
Biogenèse des CircRNA, propriétés moléculaires et voies de dégradation
(Figure : vetopsy.fr d'après Hatzimalonis et coll)
3. Selon leur origine dans le transcrit pré-ARNm, on distingue trois types principaux de circRNA.
a. Les ecircRNA (exonic circRNA) sont composés uniquement d’exons provenant du transcrit pré-ARNm.
Ils constituent la forme la plus abondante de circRNA et sont localisés principalement dans le cytoplasme, où ils participent souvent à des mécanismes de régulation post-transcriptionnelle.
b. Les EIciRNA (exon-intron circRNA) contiennent à la fois des exons et des introns.
Contrairement aux ecircRNA, ils sont majoritairement localisés dans le noyau et peuvent participer à la régulation transcriptionnelle du gène dont ils sont issus.
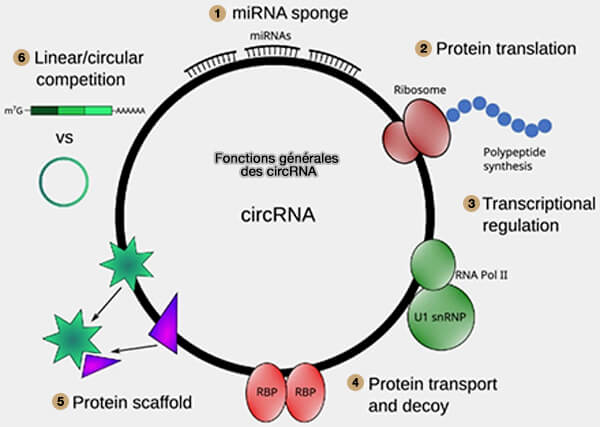
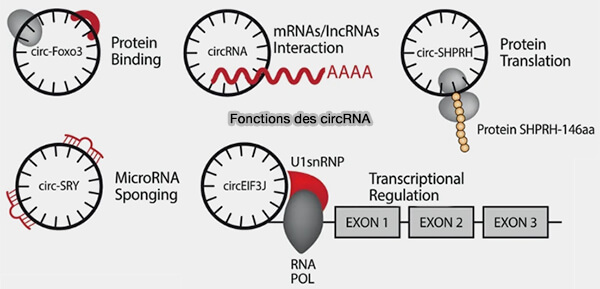
Fonctions générales des circRNA
(Figure : vetopsy.fr d'après Hatzimalonis et coll)
1. Certains circRNA agissent comme ARN compétiteurs endogènes capables de séquestrer des microARN (miRNA), i.e. miRNA sponge, limitant ainsi leur interaction avec leurs ARN messager (ARNm) cibles et modulant la répression post-transcriptionnelle exercée par ces petits ARN régulateurs.
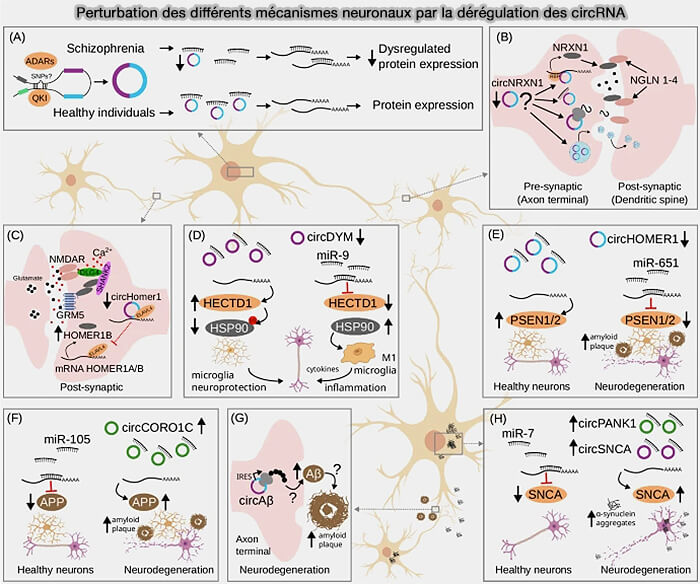
Perturbation des différents mécanismes neuronaux par la dérégulation des circRNA
(Figure : vetopsy.fr d'après Hatzimalonis et coll)
2. Bien que les circRNA soient généralement considérés comme des ARN non codants (ncRNA), certaines molécules peuvent être traduites dans des conditions particulières.
a. Ils peuvent contenir des éléments d’initiation de la traduction indépendants de la coiffe 5', tels que des des structures de type IRES (Internal Ribosome Entry Site) ou certaines modifications comme m6A, permettant le recrutement du ribosome.
Ces ARN peuvent ainsi contenir de courts cadres de lecture ouverts (ORF) capables de produire de petits peptides, mais ces événements de traduction restent rares et la grande majorité des protéines cellulaires est synthétisée à partir des ARN messager (ARNm).
3. Certains circRNA, notamment ceux localisés dans le noyau, peuvent participer à la régulation transcriptionnelle du gène dont ils sont issus en interagissant avec l’ARN polymérase II ou avec des composants du spliceosome.
4. D’autres circRNA peuvent interagir avec des protéines de liaison à l’ARN (RNA-binding proteins, RBP) et moduler leur activité, leur stabilité ou leur localisation cellulaire.
5. Certains circRNA peuvent agir comme plateformes protéiques en recrutant simultanément plusieurs protéines régulatrices.
Cette fonction d’échafaudage moléculaire favorise l’assemblage de complexes ribonucléoprotéiques et peut rapprocher des enzymes, des facteurs de régulation ou des protéines de signalisation qui agissent ensuite de manière coordonnée.
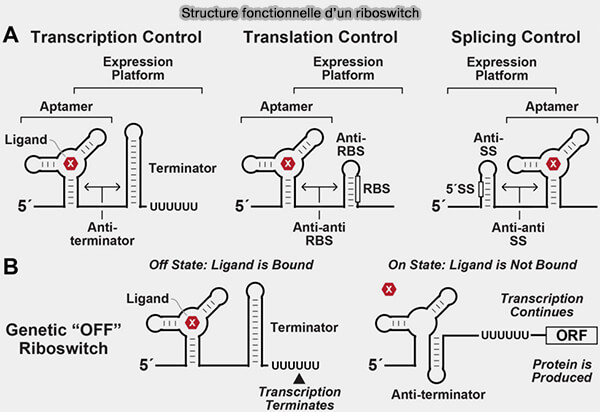
formation d’une structure terminatrice transcriptionnelle ou anti-terminatrice, contrôlant la poursuite de la transcription,
séquestration ou exposition du site de liaison du ribosome contrôlant l’initiation de la traduction,
modification de la stabilité de l’ARNm, influençant sa dégradation.
Structure fonctionnelle d'un riboswitch
(Figure : vetopsy.fr d'après Kavita et Breaker)
3. Les riboswitch sont particulièrement abondants chez les procaryotes, où ils assurent un contrôle rapide et autonome de l’expression des voies métaboliques, mais des exemples fonctionnels existent également chez les eucaryotes, notamment dans la régulation de l’épissage ou de la stabilité des ARN.
Les riboswitch les plus étudiés correspondent à des systèmes répondant à des métabolites essentiels, parmi lesquels :
les riboswitch à thiamine pyrophosphate (TPP), qui régulent les gènes impliqués dans le métabolisme de la vitamine B1 (thiamine) en modulant la transcription ou la traduction en fonction de la concentration intracellulaire de TPP,
les riboswitch à adénine, qui ajustent l’expression des gènes du métabolisme des purines en fonction de la disponibilité de l’adénine,
les riboswitch à guanine, qui régulent les voies métaboliques des purines en réponse à la concentration en guanine, souvent de manière opposée aux riboswitches à adénine,
les riboswitch à S-adénosyl méthionine (SAM), qui contrôlent l’expression de gènes impliqués dans les réactions de méthylation en fonction des niveaux de SAM,
les riboswitch à glycine, qui régulent les gènes du métabolisme de la glycine, souvent via un mécanisme coopératif impliquant deux domaines aptamères,
les riboswitch à lysine, qui régulent les gènes impliqués dans la biosynthèse, le transport ou le métabolisme de la lysine en réponse à la concentration intracellulaire de cet acide aminé





 organisation structurale et composition moléculaire)
organisation structurale et composition moléculaire)