L’euchromatine et l’hétérochromatine représentent deux états structuraux et fonctionnels de la chromatine, dont l’organisation et la dynamique sont modulées par le remodelage de la chromatine et les modifications épigénétiques.
Euchromatine
1. L'euchromatine correspond aux régions d’ADN actif, transcriptionnellement accessibles et déroulées.
Elle se situe principalement dans les régions internes du noyau au centre du nucléoplasme, bien que sa distribution puisse varier selon le type cellulaire et l’organisation du génome.
Euchromatine et hétérochromatine
(Figure : vetopsy.fr d'après Yale)
Ses histones sont ouvent enrichies en marques associées à la chromatine active, notamment l’acétylation des lysines et certaines méthylations comme H3K4me3, marquant un état ouvert favorable à la transcription.
Elle représente environ la majeure partie du génome humain, environ 92 %, valeur souvent citée mais discutable car variant selon les types cellulaires.
Remarque : l’euchromatine correspond, en microscopie électronique, aux fibrilles de chromatine diffuse et aux régions périchromatiniennes, qui apparaissent peu condensées et correspondent aux zones de chromatine transcriptionnellement active ( chromatine au microscope).
La compaction de la chromatine inhibe la transcription !
2. La condensation seule ne suffit pas à identifier l’hétérochromatine et trois critères supplémentaires doivent également être priss en compte :
une condensation différentielle persistante, supérieure à celle de la chromatine environnante, appelée hétéropycnose, observable notamment au microscope,
la méthylation de l’ADN, qui limite l’accès des facteurs de transcription,
la répression médiée par les complexes Polycomb, caractérisée notamment par la marque H3K27me3, qui maintient certains gènes dans un état silencieux mais potentiellement réversible selon le contexte cellulaire.
Les chromosomes humains 1, 9, 16 et le chromosome Y contiennent de grandes régions d’hétérochromatine constitutive, notamment dans leurs régions péricentromériques.
2. L'hétérochromatine facultative correspond à des régions du génome dont l’état de compaction et l’activité transcriptionnelle peuvent varier selon le type cellulaire, le stade de développement ou le contexte physiologique.
a. Le changement de conformation de l'hétérochromatine facultative s’effectue sous l’action de nombreux facteurs qui :
mobilisent les nucléosomes le long de la fibre chromatinienne,
aux protéines du groupe Polycomb (Pcg) qui participent à la répression transcriptionnelle en catalysant notamment la triméthylation de la lysine 27 de l’histone H3 (H3K27me3), tandis que les protéines du groupe Trithorax antagonisent cette répression et favorisent l’expression des gènes.
c. Un exemple emblématique d'hétérochromatine facultative est donné par le corpuscule de Barr chez les mammifères femelles, chromosome X, fortement condensé et transcriptionnellement silencieux par le processus de lyonisation, d'après le nom de la généticienne Mary F. Lyon (1925-2014).
L’autre chromosome X reste actif sous forme majoritairement euchromatique.
Ce mécanisme permet d’assurer la compensation de dose des gènes liés au chromosome X : chez les femelles (XX), l’inactivation d’un des deux chromosomes X empêche une double expression de ces gènes par rapport aux mâles (XY), qui ne possèdent qu’un seul chromosome X.
Remodelage de la chromatine
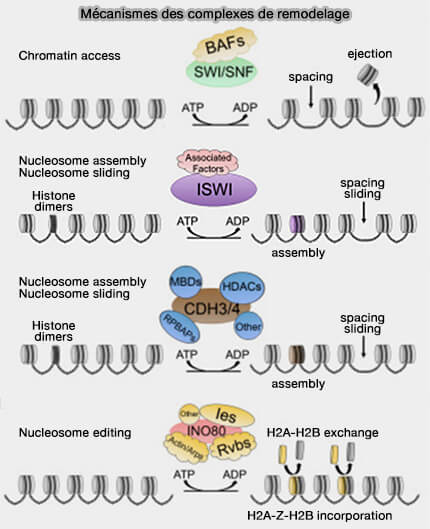
Le remodelage de la chromatine correspond à un ensemble de mécanismes permettant de modifier la position, l’organisation ou la composition des nucléosomes le long de l’ADN qui contribue à moduler localement la compaction de la chromatine pour faciliter l’accès de la machinerie impliquée dans :
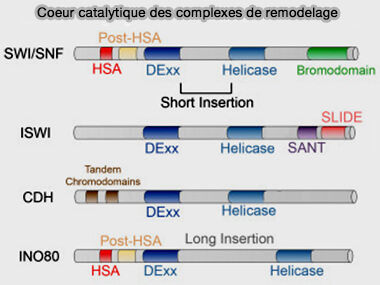
a. La famille SWI/SNF regroupe des complexes de remodelage de la chromatine caractérisés par la présence d’une ATPase de la famille SNF2, telle que BRG1 (SMARCA4) ou BRM (SMARCA2) chez les eucaryotes, dont la fonction principale est d'ouvrir la chromatine.
Ces complexes sont capables de déstabiliser les interactions entre l’ADN et les histones, entraînant le déplacement, le repositionnement ou parfois l’éjection de nucléosomes.
Mécanisme des complexes de remodelage
(Figure : vetopsy.fr d'après Magaña-Acosta et coll)
Ils jouent ainsi un rôle majeur dans l’activation transcriptionnelle, en facilitant l’accès de la machinerie transcriptionnelle aux promoteurs et aux régions régulatrices du génome.
b. La famille ISWI comprend des complexes de remodelage caractérisés par la présence d’une ATPase de type ISWI, comme SNF2H (SMARCA5) ou SNF2L (SMARCA1), dont la fonction principale est l'organisation régulière des nucléosomes, permettant la régulation transcriptionnelle et la stabilité du génome.
Ces complexes interviennent principalement dans l’organisation régulière des nucléosomes le long de l’ADN et contribuent au maintien de la structure chromatinienne.
Parmi ces complexes figure notamment NURF (Nucleosome Remodeling Factor), qui participe au repositionnement des nucléosomes et à la régulation de l’accessibilité de l’ADN et dont BPTF est la plus grande sous-unité.
Cette capacité de reconnaissance permet à ces complexes d’être recrutés sur des régions chromatiniennes spécifiques et de participer à la régulation transcriptionnelle.
Sous-unités des complexes de remodelage
(Figure : vetopsy.fr d'après Hasan et Ahuja)
d. La famille INO80, ATPase INO80, comprend des complexes de remodelage de la chromatine impliqués dans plusieurs processus essentiels du métabolisme du génome, dont la fonction principale est la modification de la composition du nucléosome (nucleosome editing).
Ces complexes sont notamment capables de remodeler les nucléosomes et de favoriser l’incorporation ou l’échange de variants d’histones, comme H2A.Z.
Par leur capacité à modifier la composition et l’organisation des nucléosomes, les complexes de la famille INO80 contribuent au maintien de l’intégrité du génome et à l’adaptation dynamique de la chromatine aux besoins fonctionnels de la cellule.
3. Le remodelage de la chromatine agit en étroite interaction avec les modifications post-traductionnelles des histones et les variants d’histones ( code des histones).
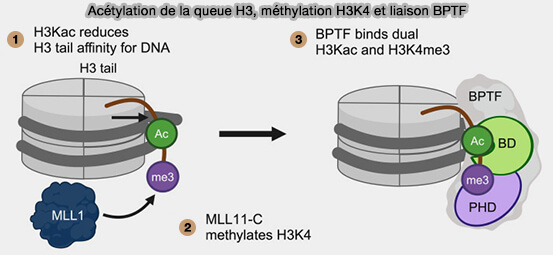
a. Certaines protéines lectrices, comme BPTF (Bromodomain PHD Finger Transcription Factor), jouent un rôle important en assurant le couplage entre la lecture des marques d’histones et le remodelage des nucléosomes.
BPTF, sous-unité majeure du complexe NURF, reconnaît notamment H3K4me3 par son domaine PHD et certaines lysines acétylées par son bromodomaine, ce qui favorise le recrutement et l’activité du complexe sur la chromatine.
Cette reconnaissance permet le repositionnement des nucléosomes et contribue ainsi à établir ou à maintenir des états chromatiniens accessibles au niveau de régions transcriptionnellement actives, notamment au niveau des gènes impliqués dans le développement, les programmes transcriptionnels embryonnaires, la régulation de l’identité cellulaire et certains gènes oncogéniques.
b. La reconnaissance de ces marques peut permettre le recrutement de complexes de remodelage, contribuant ainsi à établir ou à maintenir des états chromatiniens actifs ou répressifs.
Structure suivant le cycle cellulaire
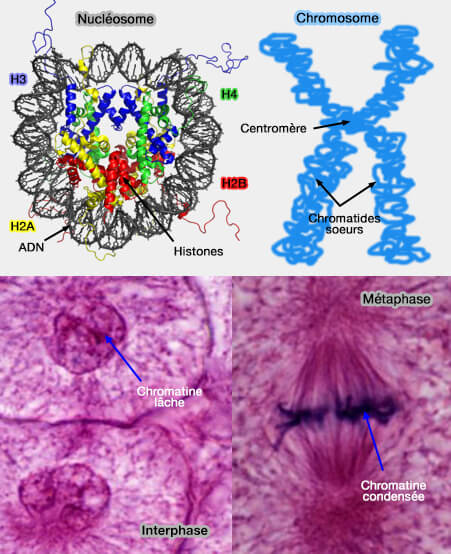
La structure de la chromatine varie pendant le cycle cellulaire.
Nucléosome et chromosome
(Figure : vetopsy.fr d'après Zephiris et Studyblue)
1. Pendant l'interphase, la chromatine est constituée d’un mélange d’euchromatine relativement accessible et d’hétérochromatine plus condensée, permettant la transcription, la réplication et la réparation de l’ADN .
Cette organisation facilite l’accès des ARN polymérases, de l’ADN polymérases et des facteurs de réparation aux régions codantes et régulatrices du génome.
La transcription est globalement arrêtée durant la mitose, et le bookmarking assure que certains gènes reprennent rapidement leur activité après la division cellulaire.




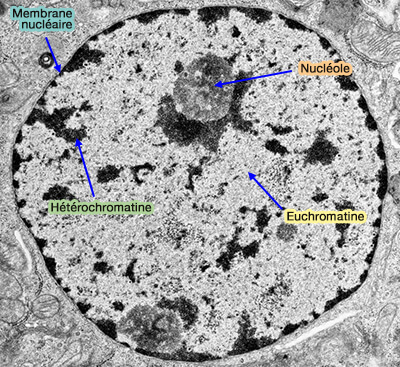
 chromatine au microscope).
chromatine au microscope).