


Acides nucléiques
ARN : dégradation
Voies et mécanismes enzymatiques
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La dégradation des ARN regroupe les mécanismes cellulaires qui contrôlent la stabilité des transcrits, régulent l’expression génique et assurent leur élimination grâce à l’action des ribonucléases et du complexe de l’exosome.
Les molécules d’ARN cellulaires possèdent une durée de vie limitée et la dégradation des ARN constitue un processus fondamental de régulation de l’expression génique, car elle permet :
- d’ajuster la quantité d’ARN présents dans la cellule,
- de contrôler leur durée de vie,
- d’éliminer les transcrits devenus inutiles ou présentant des anomalies,
- de participer à la défense cellulaire contre certains ARN viraux ou dérivés d’éléments transposables (transposons).
Chez les eucaryotes, ces mécanismes de dégradation sont répartis entre le noyau et le cytoplasme et reposent sur des ensembles distincts de ribonucléases, de complexes multiprotéiques et de voies de surveillance spécialisées.
Dégradation nucléaire et cytoplasmique des ARN
La dégradation des ARN repose sur des mécanismes partiellement distincts dans le noyau et le cytoplasme, qui assurent à la fois le renouvellement des ARN fonctionnels et l’élimination des ARN aberrants.
1. Dans le noyau, les processus de surveillance sont principalement associés :
- au contrôle qualité des transcrits,
- à la maturation des ARN (RNA processing),
- à l’élimination des ARN aberrants par l’exosome nucléaire et ses cofacteurs associés.
2. Dans le cytoplasme, la dégradation des ARN est davantage couplée :
- à la traduction,
- à la stabilité des ARN messagers (ARNm),
- aux mécanismes de régulation post-transcriptionnelle et de surveillance.

Les principales voies de dégradation cytoplasmique des ARN messagers (ARNm) sont étudiées dans un chapitre spécifique.
La surveillance des autres ARN cellulaires, notamment des ARNr, ARNt, snARN, snoARN ou différents ARN nucléaires non codants, est principalement associée à leur maturation et au contrôle qualité nucléaire assuré par le complexe de l'exosome et ses cofacteurs associés.
3. Les cellules possèdent également plusieurs mécanismes spécialisés de surveillance des ARN permettant de détecter et d'éliminer différents ARN aberrants produits au cours de la transcription, de la maturation ou de la traduction.
Les mécanismes de surveillance des ARN sont étudiés dans un chapitre spécifique.
Ribonucléases (RNases)
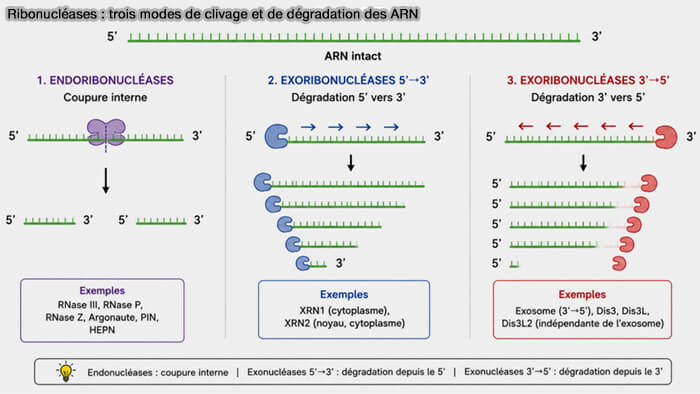
Le terme RNase (ribonucléase) désigne l’ensemble des enzymes capables de cliver, raccourcir ou dégrader les molécules d’ARN.
Ces enzymes regroupent un ensemble hétérogène appartenant à plusieurs familles structurales et fonctionnelles capables de cliver les ARN selon des mécanismes distincts.
(Figure : vetopsy.fr)
1. La dégradation des ARN repose principalement sur l’action de deux grandes catégories de ribonucléases capables de cliver les molécules d’ARN selon des mécanismes distincts :
- les endoribonucléases
- les exoribonucléases.
2. Chez les eucaryotes, ces différentes ribonucléases fonctionnent fréquemment au sein de complexes multiprotéiques spécialisés capables de coordonner la reconnaissance, le remodelage et la dégradation des ARN cibles.
L’exosome constitue notamment le principal complexe exoribonucléolytique 3′ → 5′ impliqué dans la maturation, la surveillance et la dégradation de nombreux ARN cellulaires.
La classification des ribonucléases est présentée de manière détaillée dans la page Wikipédia, et pour une analyse plus poussé même si l'étude concerne les bactéries dans Bacterial ribonucleases and their roles in RNA metabolism (2019).
Endoribonucléases
1. Les endoribonucléases réalisent des coupures internes dans les molécules d’ARN, générant des fragments plus courts.
a. Ces enzymes reconnaissent généralement :
- des séquences nucléotidiques spécifiques,
- des structures secondaires particulières,
- des régions double brin,
- certains ARN anormaux.
b. Les principales catégories d’endoribonucléases sont présentées dans le tableau suivant.
| Famille/ catégorie |
Domaine ou organisation caractéristique |
Exemples | Fonctions principales |
|---|---|---|---|
| RNase A | Domaine RNase A |
|
Dégradation des ARN extracellulaires chez les vertébrés |
| RNase T1 | Domaine RNase T1 | RNase T1 | Clivage spécifique des ARN après les résidus guanosine |
| RNase III | Domaine RNase III | Clivage des ARN double brin |
|
| RNases H | Domaine RNase H | Dégradation des hybrides ARN-ADN |
|
| RNases P | Complexe ribonucléoprotéique |
RNase P | Maturation 5′ des pré-ARNt |
| RNases Z | Métallo-β-lactamase/ β-CASP |
ELAC2 | Maturation 3′ des pré-ARNt |
| Domaine PIWI | Domaine apparenté aux RNases H |
||
| Domaine PIN | Domaine PIN | Maturation ou surveillance des ARN |
|
| Domaine HEPN | Domaine HEPN |
|
Maturation ou dégradation des ARN |
2. Les RNases III constituent une superfamille majeure spécialisée dans le clivage des ARN double brin ou des régions appariées des ARN.
a. Cette famille comprend notamment Drosha et Dicer qui jouent un rôle central dans :
3. Les RNases H dégradent spécifiquement le brin ARN des hybrides ARN-ADN et interviennent notamment dans l’élimination des amorces ARN lors la maturation des fragments d’Okazaki au cours de la réplication de l'ADN.
Certaines protéines Argonaute possèdent une activité endoribonucléase dite " slicer ", portée par un domaine PIWI apparenté aux RNases H, permettant le clivage spécifique d’un ARN cible reconnu par complémentarité avec un petit ARN guide ( biogenèse et mécanisme d’action des miRNA).
biogenèse et mécanisme d’action des miRNA).
4. Les RNases P et MRP sont des endoribonucléases ribonucléoprotéiques apparentées.
- La RNase P intervient dans la maturation des précurseurs d’ARNt en clivant leur extrémité 5′.
- La RNase MRP participe notamment au clivage de certains intermédiaires des pré-ARNr au cours de la ribogenèse.
5. Les RNases Z, également appelées tRNase Z, participent à la maturation des précurseurs d’ARNt en clivant leur extrémité 3′ afin de générer l’extrémité qui recevra ultérieurement la séquence CCA chez de nombreux organismes.
6. Les endoribonucléases à domaine PIN (PilT N-terminus) interviennent dans diverses étapes du métabolisme des ARN, notamment leur maturation, leur surveillance et leur dégradation.
Cette famille comprend notamment :
- NOB1, impliquée dans la maturation de l’ARNr 18S,
- UTP24, impliquée dans la maturation de l’ARNr 45S,
- SMG6, qui participe à la dégradation des ARNm ciblés par la voie NMD (nonsense-mediated decay).
7. Les endoribonucléases à domaine HEPN interviennent dans différents mécanismes de maturation et de contrôle qualité des ARN.
Cette famille comprend notamment :
- LAS1, impliquée dans la maturation des précurseurs d’ARNr,
- IRE1α, dont l’activité endoribonucléase participe à la réponse aux protéines mal repliées par l’épissage non conventionnel de l’ARNm XBP1 (réponse UPR du réticulum endoplasmique).
Exoribonucléases
1. Les exoribonucléases dégradent progressivement les ARN à partir de leurs extrémités libres par retrait successif de nucléotides.
a. Selon les enzymes impliquées, cette dégradation peut se produire dans deux directions principales :
- dans le sens 5′ → 3′,
- dans le sens 3′ → 5′.
b. Les principales catégories d’exoribonucléases sont présentées dans le tableau suivant.
| Famille/ catégorie |
Domaine ou organisation caractéristique |
Exemples | Fonctions principales |
|---|---|---|---|
| RNase II/R | Domaine RNB | Dégradation, maturation et surveillance des ARN |
|
| Exosome | Complexe multiprotéique associé à des exoribonucléases |
Dégradation et maturation des ARN nucléaires et cytoplasmiques |
|
| XRN | Domaine XRN | Dégradation 5′ → 3′ des ARN |
|
| DEDD | Domaine DEDD |
|
Maturation et surveillance des ARN |
| PAN2-PAN3 | Exoribonucléase de type déadénylase |
|
Déadénylation initiale des ARNm |
| CCR4-NOT | Complexe de déadénylation |
|
Déadénylation des ARNm |
2. Les exoribonucléases de la famille RNase II/R correspondent principalement à des exoribonucléases 3′ ➞ 5′ conservées des bactéries aux eucaryotes.
Chez les eucaryotes, cette famille comprend notamment Dis3, Dis3L, Dis3L2.
- Contrairement à Dis3, présente à la fois dans le noyau et le cytoplasme mais principalement associée à l’exosome nucléaire, et à Dis3L, principale sous-unité catalytique de l’exosome cytoplasmique, Dis3L2 agit indépendamment de ce complexe et participe notamment à la dégradation d’ARN portant des extensions riches en uridine à leur extrémité 3'.
- Ces enzymes participent à la dégradation, à la maturation et à la surveillance de nombreux ARN cellulaires, notamment certains ARN ribosomiques (ARNr), des snARN (small nuclear RNA) et des ARN messagers (ARNm) aberrants.
3. Les exoribonucléases XRN correspondent à des exoribonucléases 5′ ➞ 3′ hautement processives.
- Chez les eucaryotes, XRN1 agit principalement dans le cytoplasme tandis que XRN2 agit principalement dans le noyau.
- Ces enzymes jouent un rôle central dans la dégradation des ARN décapés ainsi que dans certains mécanismes de terminaison transcriptionnelle, en dégradant l’ARN résiduel situé en aval du site de clivage du pré-ARNm.
4. Les exoribonucléases de la famille DEDD possèdent un domaine catalytique caractérisé par un motif conservé Asp-Glu-Asp-Asp (DEDD) et comprennent notamment PARN et ISG20L2, impliquées dans la maturation et la surveillance de nombreux ARN cellulaires.
Complexe de l’exosome
Le complexe de l’exosome est un grand complexe multiprotéique possédant principalement une activité exoribonucléase 3′ ➞ 5′ impliquée dans la dégradation et la maturation de nombreux ARN cellulaires.

Vous pouvez lire : The RNA exosome and RNA exosome-linked disease (2018).
Remarque : le complexe de l’exosome impliqué dans la maturation, la surveillance et la dégradation des ARN ne doit pas être confondu avec les exosomes extracellulaires, petites vésicules membranaires du système endomembranaire impliquées dans la communication intercellulaire et le transport des protéines, des lipides et des acides nucléiques.
Vue d'ensemble
1. Chez les eucaryotes, l’exosome est présent à la fois dans le noyau et dans le cytoplasme où il participe au contrôle de la stabilité et de la qualité des ARN.
a. Dans le cytoplasme, il intervient dans de nombreux processus du métabolisme des ARN, notamment :
- la dégradation des ARN messagers et de différents ARN cytoplasmiques ciblés par des mécanismes de régulation post-transcriptionnelle,
- la surveillance de la qualité des ARN et l’élimination d’ARN aberrants, incomplets ou mal maturés ayant échappé aux contrôles nucléaires.
b. Dans le noyau, l’exosome participe notamment à l’élimination de nombreux transcrits nucléaires instables ainsi qu’à la maturation de certains ARN ribosomiques (ARNr), de snARN (small nuclear RNA) et de snoARN (small nucleolar RNA).
(Figure : vetopsy.fr d'après Morton et coll)
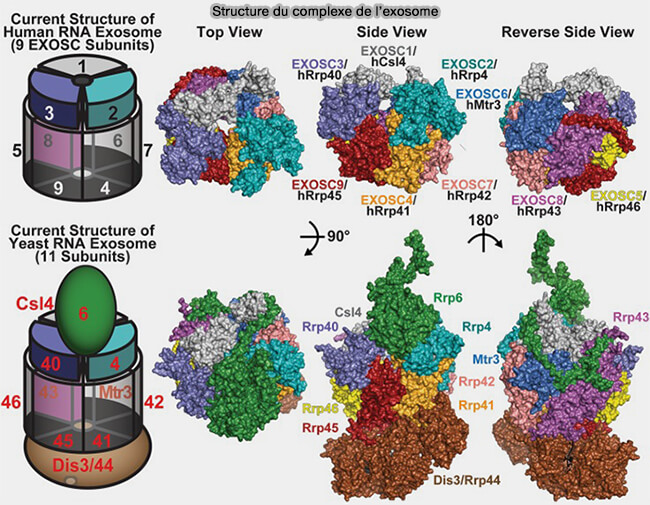
2. La structure de l'exosome est complexe.
a. L’exosome eucaryote complet (holo-exosome) comprend :
- un cœur central non catalytique (core exosome ou EXO-9) formé des sous-unités structurales organisées en anneau,
- des sous-unités catalytiques associées.
b. Différents cofacteurs de recrutement et d’activation peuvent également s’associer au complexe et participer à la reconnaissance, au remodelage et à l’acheminement des ARN vers l’exosome.
Holo-exosome
1. Le cœur central comprend :
- un hexamère de protéines de type RNase PH (EXOSC4/Rrp41, EXOSC5/Rrp46, EXOSC6/Mtr3, EXOSC7/Rrp42, EXOSC8/Rrp43 et EXOSC9/Rrp45), dérivées structuralement des ribonucléases phosphorolytiques ancestrales de type RNase PH et formant l’anneau central de l’exosome,
- surmonté d’un cap protéique (complexe cap), trimère de protéines constitué de EXOSC1/Csl4, EXOSC2/Rrp4 et EXOSC3/Rrp40 possédant des domaines de liaison aux ARN.
L’ensemble forme un canal central étroit permettant le recrutement et le guidage des ARN vers les sous-unités catalytiques associées au complexe.
- Contrairement aux exosomes bactériens et archéens, le cœur central de l’exosome eucaryote possède essentiellement un rôle structural et de canalisation des substrats ARN, la majorité de l’activité catalytique étant portée par des ribonucléases associées.
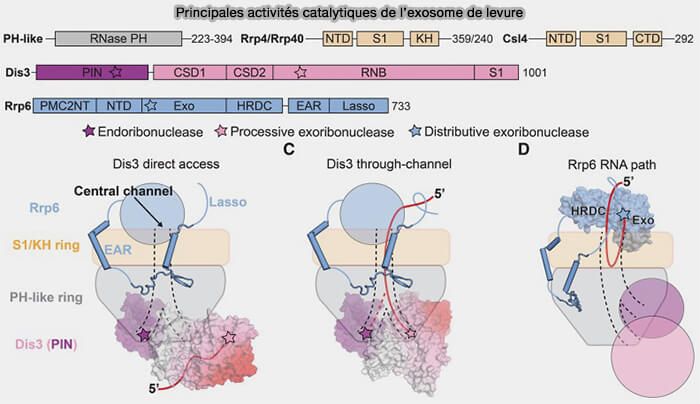
(Figure : vetopsy.fr d'après Zinder et Lima)
2. Les principales sous-unités catalytiques associées à l’exosome sont localisées à la périphérie du cœur central du complexe (Targeting RNA for processing or destruction by the eukaryotic RNA exosome and its cofactors 2017).
a. Dis3 (EXOSC11), une exoribonucléase processive 3′ → 5′ de la famille RNase II/R possédant également un domaine endoribonucléase PIN, est associée à la face inférieure de l’anneau de l’exosome.
- Les ARN engagés dans le canal central du complexe peuvent être transférés vers le site catalytique de Dis3, qui assure alors une dégradation processive extensive du substrat ARN.
- Le domaine PIN de Dis3 permet également des clivages endoribonucléolytiques internes sur certains ARN structurés, aberrants ou fortement associés à des complexes ribonucléoprotéiques.
b. XOSC10/Rrp6, une exoribonucléase distributive principalement impliquée dans les fonctions nucléaires de maturation et de surveillance des ARN, est localisée au niveau de la face supérieure du complexe, à proximité du cap protéique de liaison aux ARN.
- Contrairement à Dis3, EXOSC10 agit généralement de manière plus limitée et distributive en retirant progressivement quelques nucléotides à partir des extrémités 3′ accessibles avant de se dissocier du substrat.
- EXOSC10 participe ainsi au raccourcissement (trimming) et à la maturation de nombreux ARN nucléaires, mais également au contrôle qualité et à l’élimination de différents ARN mal maturés ou instables.
c. Un même ARN peut être successivement pris en charge par EXOSC10, puis transféré vers Dis3 selon son état de maturation, sa structure ou son degré d’anomalie, illustrant la coopération fonctionnelle entre les différentes activités catalytiques associées à l’exosome eucaryote.
3. L’organisation générale de l'holo-exosome est globalement conservée entre le noyau et le cytoplasme, mais certaines différences existent au niveau des sous-unités catalytiques associées au complexe.
- Dans le noyau, l’exosome est principalement associé aux deux exoribonucléases précédentes, i.e. Dis3 (EXOSC11) et EXOSC10/Rrp6.
- Dans le cytoplasme, l’exosome est principalement associé à une seule exoribonucléase, Dis3L, paralogue cytoplasmique de Dis3 (EXOSC11).
Facteurs associés
Les ARN cibles destinés à la dégradation, ARN aberrants, mal maturés, instables ou présentant des défauts de repliement, sont reconnus par différents cofacteurs de recrutement et complexes adaptateurs qui dirigent les substrats vers l’exosome.
Les cofacteurs associés à l’exosome diffèrent partiellement entre le noyau et le cytoplasme, ce qui contribue à orienter le complexe vers des populations distinctes d’ARN cibles.
Facteurs associés dans le noyau
Dans le noyau, plusieurs complexes adaptateurs et cofacteurs spécialisés participent au recrutement des ARN cibles vers l’exosome nucléaire.
1. Chez la levure, le complexe TRAMP (Trf4/5-Air1/2-Mtr4 polyadenylation complex), qui agit comme une plateforme de surveillance nucléaire couplant reconnaissance des ARN aberrants, oligoadénylation et recrutement fonctionnel de l’exosome nucléaire, joue un rôle majeur dans cette reconnaissance.
a. Il comprend plusieurs protéines.
- les poly(A) polymérases non canoniques Trf4 ou Trf5 ajoutent de courtes queues oligoadénylées à différents ARN nucléaires aberrants, mal maturés ou excédentaires, créant ainsi un signal favorisant leur reconnaissance et leur adressage vers l’exosome nucléaire.
- les protéines de liaison aux ARN Air1 ou Air2 possèdent plusieurs domaines à doigts de zinc impliqués dans la reconnaissance et le recrutement de différents substrats ARN nucléaires.
- l’hélicase ARN Mtr4, une hélicase de la famille Ski2-like associée directement à l’exosome nucléaire, déroule les structures secondaires des ARN et facilite leur engagement ainsi que leur translocation vers le canal central du complexe exosomique.
b. Le complexe TRAMP interagit principalement avec la face nucléaire supérieure de l’exosome, au niveau du cap formé par les sous-unités EXOSC1/2/3 et surtout via l’association fonctionnelle de Mtr4 avec EXOSC10/Rrp6.
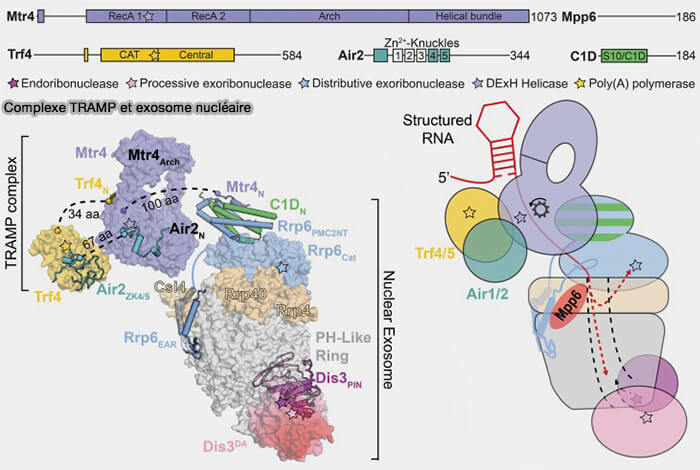
(Figure : vetopsy.fr d'après Zinder et Lima)
2. Chez les mammifères, il n’existe pas de complexe TRAMP strictement équivalent à celui décrit chez la levure.
a. Plusieurs complexes adaptateurs nucléaires spécialisés assurent des fonctions analogues de surveillance des ARN et de recrutement vers l’exosome nucléaire.
- Le complexe NEXT (Nuclear EXosome Targeting) intervient principalement dans la surveillance de nombreux ARN nucléaires courts, instables ou rapidement dégradés, notamment certains PROMPTs, eRNA, snARN ou transcrits issus de régions intergéniques.
- Le complexe PAXT (Poly(A) tail EXosome Targeting) cible davantage des ARN nucléaires polyadénylés plus longs, notamment certains lncRNA, ARN mal maturés ou transcrits retenus dans le noyau.
b. Ces complexes recrutent l’hélicase ARN MTR4, qui assure le transfert des ARN cibles vers l’exosome nucléaire pour leur maturation ou leur dégradation.
Facteurs associés dans le cytoplasme
Dans le cytoplasme, l’exosome s’associe à plusieurs cofacteurs impliqués dans la dégradation des ARN messagers.
1. Le complexe SKI (Superkiller complex) joue un rôle central dans plusieurs mécanismes de surveillance et de dégradation 3′ ➞ 5′ des ARN messagers cytoplasmiques et comprend notamment :
- l’hélicase ARN Ski2 (SKIV2L chez les eucaryotes supérieurs), une hélicase de la famille Ski2-like localisée au centre fonctionnel du complexe, qui utilise l’hydrolyse de l’ATP pour dérouler les structures secondaires des ARN et favoriser leur translocation vers le canal central de l’exosome,
- les protéines Ski3 et Ski8, qui forment une plateforme structurale stabilisant l’organisation du complexe SKI et participant à son interaction avec l’exosome cytoplasmique,
- la protéine Ski7, un facteur adaptateur associé à l’exosome cytoplasmique chez la levure, impliqué dans le recrutement fonctionnel du complexe SKI vers les ARN en cours de traduction ou engagés dans des mécanismes de surveillance traductionnelle.
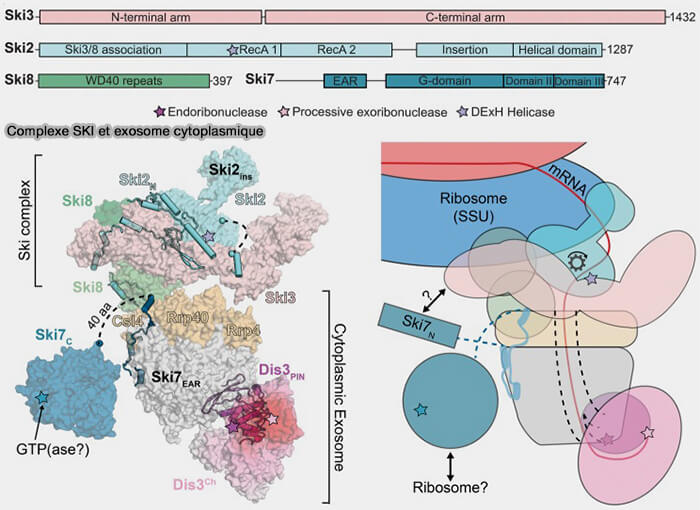
(Figure : vetopsy.fr d'après Zinder et Lima)
2. Le complexe SKI établit une interface fonctionnelle entre les ribosomes cytoplasmiques et l’exosome lors de plusieurs mécanismes de surveillance traductionnelle, notamment les voies NSD (non-stop decay) et NGD (no-go decay), ainsi que dans certaines étapes de dégradation associées à la voie NMD (nonsense-mediated decay).
Il peut notamment être recruté au niveau de ribosomes bloqués ou engagés dans des collisions ribosomiques afin de favoriser l’extraction, le déroulement puis la dégradation 3′ → 5′ des ARN messagers ciblés par l’exosome cytoplasmique.
Les ARN engagés dans le complexe SKI traversent ensuite le canal central de l’exosome avant d’être dégradés progressivement par les sous-unités catalytiques associées, principalement Dis3 dans le cytoplasme.
Principales voies de dégradation cytoplasmique des ARNm
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation