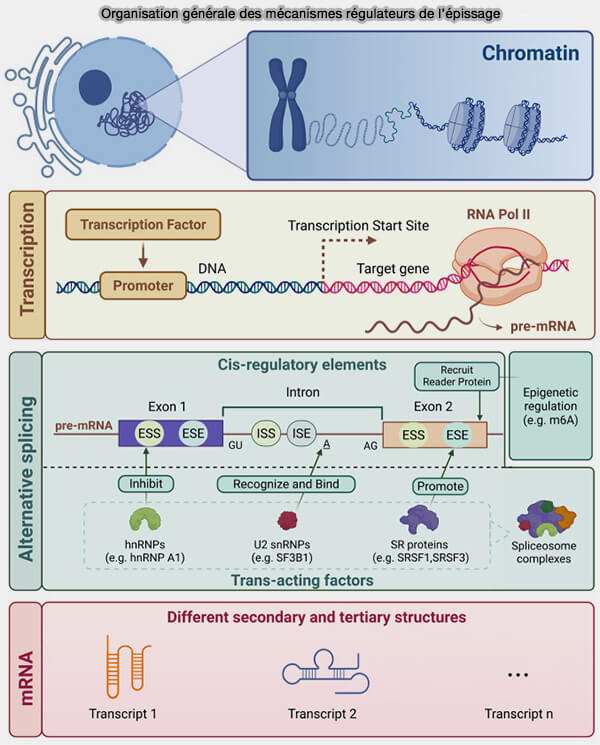
Les facteurs d’épissage régulent la reconnaissance des sites d’épissage et l’épissage alternatif des pré-ARNm, permettant la production d’isoformes d’ARNm et de protéines différentes à partir d’un même gène.
Vue d'ensemble des facteurs d'épissage
Les facteurs d’épissage (splicing factors) regroupent un ensemble de protéines et de complexes ribonucléoprotéiques régulateurs contrôlant :
Les facteurs trans correspondent à des protéines ou complexes ribonucléoprotéiques capables de reconnaître ces séquences et de moduler l’assemblage ou l’activité du spliceosome.
Les éléments cis correspondent à des séquences présentes directement dans le pré-ARNm, telles que les sites d’épissage, les ESE, ESS, ISE ou ISS.
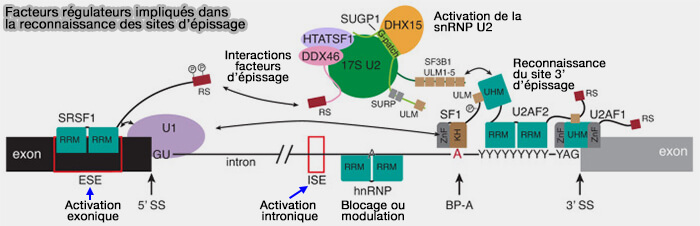
Facteurs régulateurs impliqués dans la reconnaissance des sites d’épissage
(Figure : vetopsy.fr d'après Tholen)
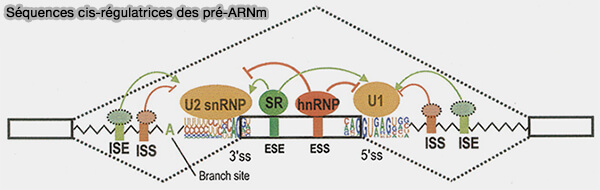
1. Les ESE (exonic splicing enhancers) correspondent à des séquences activatrices localisées dans les exons.
Ils recrutent fréquemment des protéines SR favorisant l’assemblage du spliceosome et l’inclusion des exons voisins dans l’ARNm mature.
Séquences cis-régulatrices des pré-ARNm
(Figure : vetopsy.fr d'après Wang et Burge)
2. Les ESS (exonic splicing silencers) correspondent à des séquences inhibitrices localisées dans les exons.
Ils recrutent souvent des protéines hnRNP capables d’inhiber localement le recrutement du spliceosome ou de favoriser l’exclusion de certains exons.
3. Les ISE (intronic splicing enhancers) sont des séquences activatrices situées dans les introns et participant à la reconnaissance de certains sites d’épissage.
4. Les ISS (intronic splicing silencers) correspondent à des séquences introniques inhibitrices capables de réprimer l’utilisation de certains sites d’épissage ou de moduler la sélection des exons.
Facteurs protéiques régulateurs d'épissage
Les principaux facteurs protéiques d’épissage impliqués dans l’épissage spliceosomal comprennent notamment :
les protéines SR impliquées dans l’activation de l’épissage et la reconnaissance des exons,
les protéines hnRNP intervenant dans la répression ou la modulation de certains sites d’épissage,
différents facteurs auxiliaires spécialisés contrôlant l’assemblage du spliceosome, la reconnaissance des séquences consensus et la régulation tissu-spécifique de l’épissage.
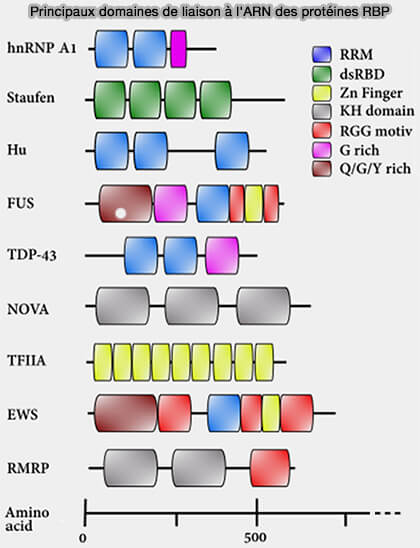
Domaines de liaison à l’ARN et protéines (RBP)
1. Les facteurs protéiques régulateurs de l’épissage appartiennent à une vaste famille de protéines de liaison à l’ARN (RNA-binding proteins ou RBP) impliquées dans de nombreux processus du métabolisme des ARN, notamment (From Structure to Function of RNA-Binding Domains 2011) :
Principaux domaines de liaison à l'ARN des protéines RBP
(Figure : vetopsy.fr d'après Ocharán-Mercado et coll)
les protéines de stabilité et de dégradation des ARNm comme HuR/ELAVL1, TTP/ZFP36, AUF1/HNRNPD ou KSRP/KHSRP,
Certains motifs interagissent principalement avec des ARN simple brin, comme les domaines RRM ou KH fréquemment retrouvés dans les facteurs d’épissage,
d’autres reconnaissent préférentiellement des ARN double brin ou des structures tridimensionnelles plus complexes comme les motifs dsRBD, PUF, PAZ, PIWI, S1 ou La motif…
Remarque : les hélicases ARN-dépendantes de la famille DEAD-box constituent quant à elles une classe particulière de protéines capables de remodeler dynamiquement les interactions ARN-ARN et ARN-protéines au sein des complexes ribonucléoprotéiques.
Ces interactions contribuent notamment à la reconnaissance des sites d’épissage, au recrutement du spliceosome et à la régulation de l’épissage alternatif.
Les trois domaines classiques des RBD
(Figure : vetopsy.fr d'après Fang et coll)
a. Le motif RRM (RNA Recognition Motif) constitue le domaine de liaison à l’ARN le plus fréquent chez les protéines d’épissage.
Ce domaine d’environ 80-90 acides aminés adopte généralement une structure β1α1β2β3α2β4 formant une surface de liaison capable d’interagir avec des ARN simple brin par des interactions électrostatiques, hydrophobes et des empilements aromatiques avec les bases azotées.
Les protéines SR, de nombreuses hnRNP ainsi que plusieurs facteurs d’épissage alternatifs possèdent un ou plusieurs domaines RRM impliqués dans la reconnaissance des séquences ESE, ESS, ISE ou ISS des pré-ARNm.
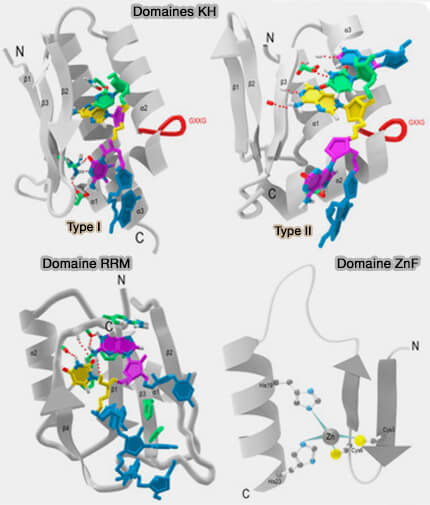
b. Les domaines KH (K Homology domain) correspondent à une autre famille importante de motifs de liaison à l’ARN présents dans certains facteurs régulateurs de l’épissage.
Ces domaines reconnaissent principalement des ARN simple brin grâce à une poche de liaison impliquant notamment un motif protéique consensus conservé de type GXXG riche en glycine.
Des protéines neuronales telles que NOVA utilisent notamment des domaines KH, tandis que certains facteurs RBFOX possèdent principalement des domaines RRM permettant de reconnaître des motifs spécifiques impliqués dans la régulation tissu-spécifique de l’épissage alternatif.
c. Certains facteurs d’épissage possèdent également des domaines à doigt de zinc (zinc finger) ou d’autres motifs spécialisés capables de stabiliser les interactions ARN-protéines ou ARN-ARN au sein des complexes spliceosomaux.
Leur fixation favorise généralement le recrutement du spliceosome au niveau des jonctions exon-intron voisines et stimule l’inclusion des exons dans l’ARNm mature.
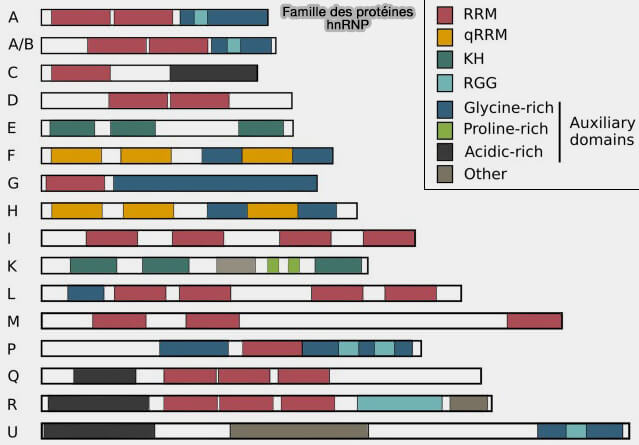
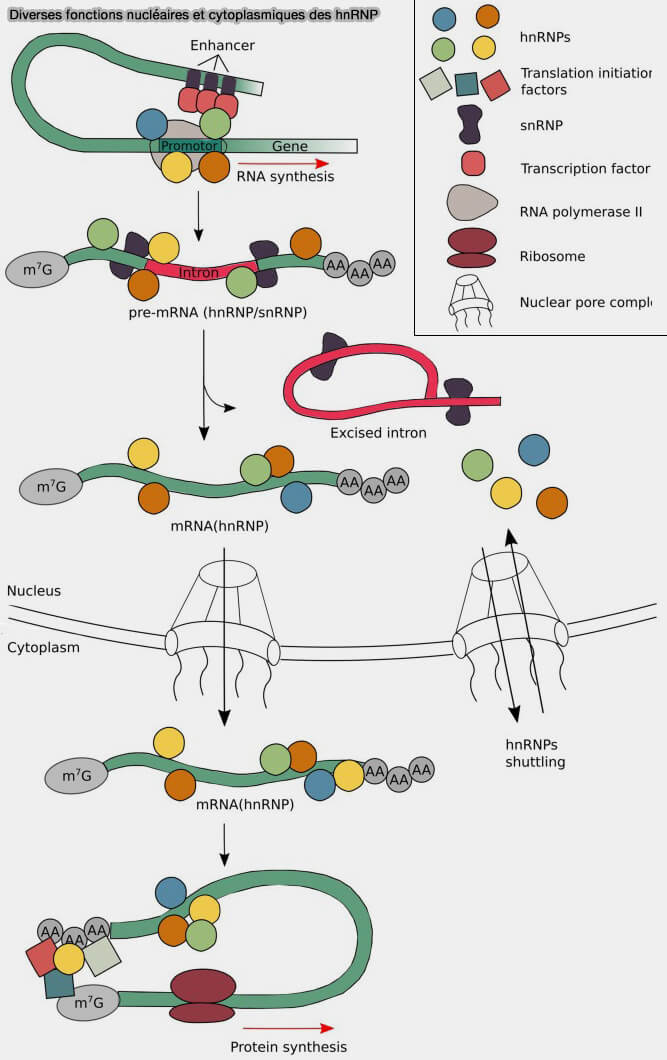
Structure des protéines hnRNP
(figure : vetopsy.fr d'après Geuens et coll)
Elles comprennent de nombreux facteurs tels que hnRNPA1, hnRNPC, hnRNPH, hnRNPK ou PTBP1 (polypyrimidine tract-binding protein 1), capables de reconnaître différentes séquences régulatrices des pré-ARNm et de moduler la sélection des sites d’épissage.
Dans l’épissage spliceosomal, les protéines hnRNP reconnaissent fréquemment des séquences régulatrices inhibitrices et antagonisent souvent l’action des protéines SR.
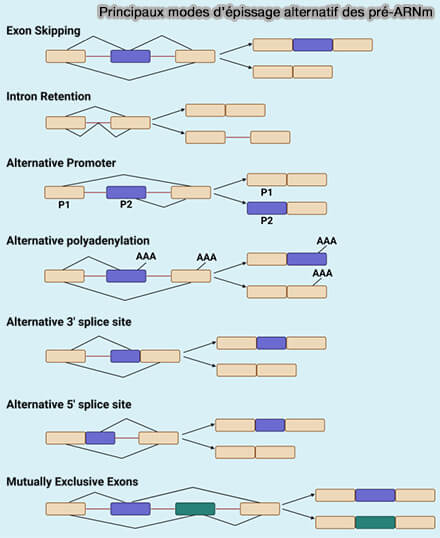
1. L’épissage alternatif correspond à un mécanisme de maturation des pré-ARNm permettant de produire plusieurs isoformes d’ARNm à partir d’un même gène grâce à l’utilisation différentielle des sites d’épissage.
b. L’épissage alternatif est particulièrement abondant chez les organismes multicellulaires complexes et concerne la majorité des gènes humains codant des protéines.
le saut d’exon (exon skipping), correspondant à l’inclusion ou l’exclusion d’un exon entier,
l’utilisation alternative de sites donneurs 5′ d’épissage,
l’utilisation alternative de sites accepteurs 3′ d’épissage,
les exons mutuellement exclusifs, dans lesquels un seul exon parmi plusieurs possibilités est retenu dans l’ARNm mature,
la rétention d’intron (intron retention), correspondant à la conservation partielle ou complète d’un intron dans le transcrit mature.
Remarque : certains mécanismes d’épissage non canonique peuvent également conduire à la formation d’ARN circulaires (circRNA) par un mécanisme de rétro-épissage (back-splicing), dans lequel un site donneur d’épissage situé en aval est relié à un site accepteur localisé en amont ( circRNA).
3. La régulation de l’épissage alternatif dépend de l’action coordonnée de nombreux facteurs d’épissage et des séquences régulatrices contenues dans les pré-ARNm.
Les protéines SR favorisent généralement l’utilisation de certains sites d’épissage et l’inclusion des exons,
Plusieurs protéines hnRNP inhibent localement certains événements d’épissage ou favorisent l’exclusion exonique.
Remarque : la sélection des sites d’épissage dépend également de la force des séquences consensus reconnues par le spliceosome, de l’organisation structurale du pré-ARNm, de l’accessibilité des séquences régulatrices et du contexte transcriptionnel et chromatinien au cours de la transcription.
a. Parmi les fonctions associées à l’épissage alternatif, on peut notamment citer :
celui du récepteur NMDA qui peut modifier la composition de certaines sous-unités synaptiques et influencer les propriétés de transmission et de plasticité neuronales,
celui du gène de la tropomyosine qui produit différentes isoformes adaptées soit aux cellules musculaires, soit aux cellules non musculaires, modifiant les interactions avec les filaments d’actine et les propriétés du cytosquelette.
b. Plusieurs facteurs régulateurs participent au contrôle de l’épissage alternatif.
Les protéines RBFOX ((RBFOX1, RBFOX2 et RBFOX3) et NOVA (NOVA1 et NOVA2) jouent notamment un rôle majeur dans la régulation de nombreux événements d’épissage tissu-spécifiques, en particulier dans le système nerveux.
PTBP1 participe quant à elle au contrôle de nombreux programmes d’épissage associés aux cellules progénitrices et à la différenciation neuronale.
Les protéines MBNL (MBNL1, MBNL2 et MBNL3) interviennent notamment dans la maturation des profils d’épissage musculaires et développementaux.
Dans certains cas, l’épissage alternatif peut également modifier les régions non codantes des ARNm ou entraîner l’apparition de codons stop prématurés, influençant ainsi la stabilité des transcrits, leur traduction ou leur élimination par les mécanismes de surveillance des ARN tels que la voie NMD (nonsense-mediated decay).