Le spliceosome est une machinerie ribonucléoprotéique dynamique assurant l’épissage des pré-ARNm grâce à l’action coordonnée des snARN, des snRNP et du centre catalytique ribozymique.
L’activité du spliceosome repose sur des réarrangements structuraux successifs permettant l’assemblage du centre catalytique et la réalisation des réactions de transestérification. ( centre catalytique).
Chez les eucaryotes, deux formes apparentées de spliceosome coexistent :
le spliceosome majeur, responsable de l’épissage de plus de 99 % des introns,
le spliceosome mineur, spécialisé dans l’excision d’une faible population d’introns rares appelés introns U12.
Structure générale du spliceosome majeur
snRNP majeures du spliceosome
Les snRNP (small nuclear ribonucleoproteins) constituent les unités fonctionnelles fondamentales du spliceosome.
1. Le spliceosome majeur responsable de la majorité des réactions d’épissage des pré-ARNm eucaryotes repose principalement sur les snRNP spliceosomales.
2. Chaque snRNP spliceosomale possède une composition spécifique associant un snARN, un cœur protéique commun et des protéines spécialisées (rôle des principales snRNP spliceosomales).
U1-70K et U1C participent à la reconnaissance et à la stabilisation du site donneur d’épissage 5′.
U1A contribue à l’assemblage et à la stabilité de la particule.
b. La snRNP U2 est constituée du snARN U2, des protéines Sm ainsi que des complexes protéiques SF3A, composé des sous-unités SF3A1, SF3A2 et SF3A3, et SF3B, composé notamment de SF3B1, SF3B2, SF3B3, SF3B4, SF3B5, SF3B6 et SF3B7.
Les complexes SF3A et SF3B stabilisent l’interaction de U2 avec le point de branchement, SF3B1 jouant un rôle central dans la reconnaissance de cette région.
Le snARN U4 est associé aux protéines Sm et interagit notamment avec les protéines PRPF3, PRPF4, PRPF31 et CYPH/PPIH, qui participent à l’assemblage et à la stabilisation du complexe U4/U6.
le snARN U6 est associé à plusieurs protéines LSm, impliquées dans sa stabilité, sa maturation et les réarrangements structuraux conduisant à l'activation du spliceosome.
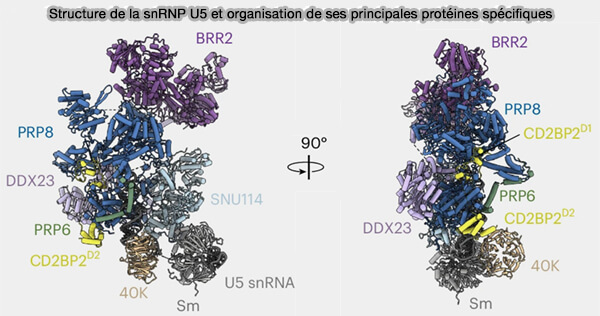
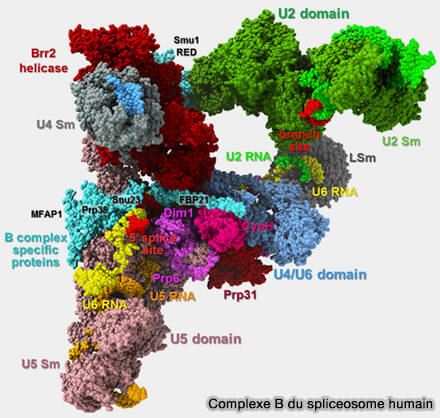
Structure de la snRNP U5 et organisation de ses principales protéines spécifiques
(Figure : vetopsy.fr d'après Doggett et coll)
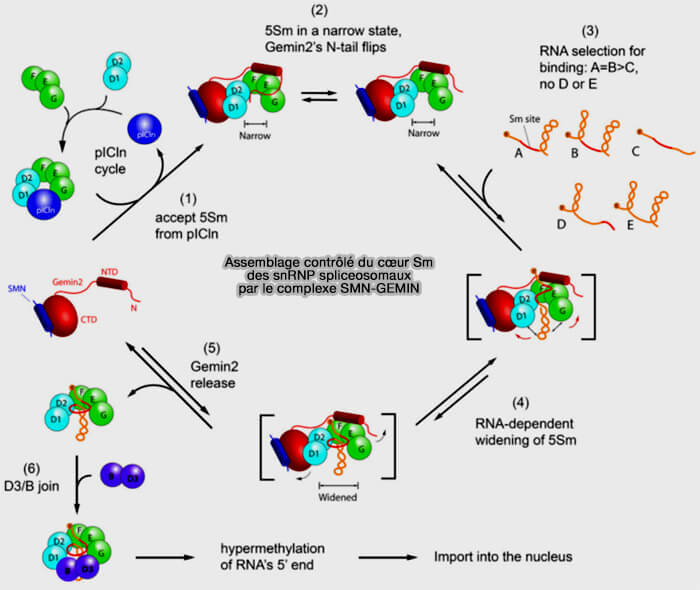
Le site Sm sert de plateforme d’assemblage permettant la fixation coordonnée des protéines Sm par des interactions spécifiques ARN-protéines et protéines-protéines conduisant à la formation de l’anneau Sm caractéristique des snRNP spliceosomales
Assemblage du cœur SM des snRNP
(Figure : vetopsy.fr d'après Yi et coll)
Cet anneau constitue le cœur structural des snRNP spliceosomales majeures U1, U2, U4 et U5 et participe à :
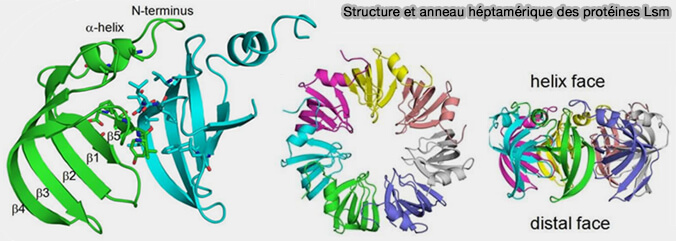
Structure et anneau héptamérique des protéines Lsm
(Figure : vetopsy.fr d'après Moll et coll)
2. Les protéines LSm (Like-Sm) appartiennent à une famille apparentée structuralement aux protéines Sm, mais forment des complexes distincts associés à différents ARN nucléaires impliqués dans l’épissage, la maturation et la dégradation des ARN (The Lsm Proteins: Ring Architectures for RNA Capture 2011).
Dans le spliceosome, le snARN U6 ne possède pas de site Sm classique, s’associe à un anneau LSM2-8 remplaçant l’anneau Sm conventionnel et reste nucléaire.
Remarque 2 : le snRNP U7 impliqué dans la maturation des pré-ARNm d’histones réplicatifs reste aussi nucléaire et possède également un cœur Sm atypique dans lequel les protéines SmD1 et SmD2 sont remplacées par les protéines LSm10 et LSm11.
3. On note une autre différence entre les snARN.
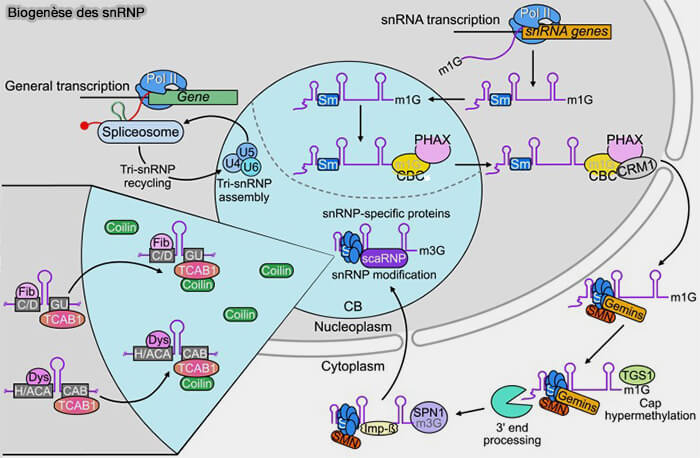
Les snARN U1, U2, U4 et U5 sont transcrits par l’ARN polymérase II, puis exportés transitoirement dans le cytoplasme où ils s’associent au complexe SMN-GEMIN et aux protéines Sm avant leur réimportation nucléaire.
Le snARN U6 constitue une exception car il est transcrit par l’ARN polymérase III.
Rôle des snRNP et centre catalytique du spliceosome majeur
Les snARN du spliceosome assurent plusieurs fonctions essentielles impliquées dans la reconnaissance des introns, l’organisation structurale du spliceosome et la catalyse des réactions d’épissage participant notamment :
à la reconnaissance des sites d’épissage,
à l’appariement avec le pré-ARNm,
à l’organisation du centre catalytique,
à la catalyse ribozymique des réactions de transestérification.
Rôle des principales snRNP spliceosomales
Chaque snRNP spliceosomale possède une organisation et des fonctions spécifiques intervenant dans différentes étapes du mécanisme spliceosomal.
1. La snRNP U1 intervient précocement lors de l’assemblage du spliceosome et assure principalement la reconnaissance initiale du site donneur 5′ d’épissage (5′SS) par appariement complémentaire entre le snARN U1 et la séquence consensus du pré-ARNm.
Cette reconnaissance contribue au positionnement initial de l’intron et à la sélection correcte des jonctions exon-intron.
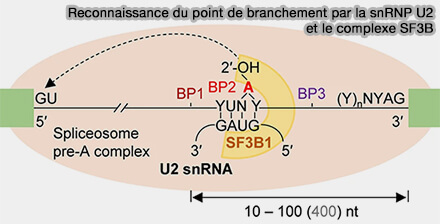
Reconnaissance du point de branchement
(Figure : vetopsy.fr d'après Xie et coll)
a. Cet appariement laisse l’adénosine du point de branchement non appariée et exposée vers le centre catalytique sous forme d’une adénosine dépariée et extrudée (bulged), configuration essentielle à la première réaction de transestérification.
La stabilisation de cette interaction implique notamment plusieurs protéines du complexe SF3B associé à la snRNP U2, en particulier SF3B1, qui participe à la reconnaissance du point de branchement et au maintien de l’adénosine catalytique dans une configuration compatible avec la formation du lariat intronique.
b. Selon le contexte de séquence, les facteurs d’épissage recrutés ou le type cellulaire, différents points de branchement (BP) peuvent être utilisés préférentiellement, ce qui peut influencer l’efficacité de l’épissage ou la sélection des sites d’épissage alternatifs.
Remarque : certaines hélicases ARN-dépendantes de la famille DExD/H-box, telles que DHX15, participent également au remodelage et au contrôle qualité des interactions entre la snRNP U2 et le point de branchement au cours de l’assemblage spliceosomal.
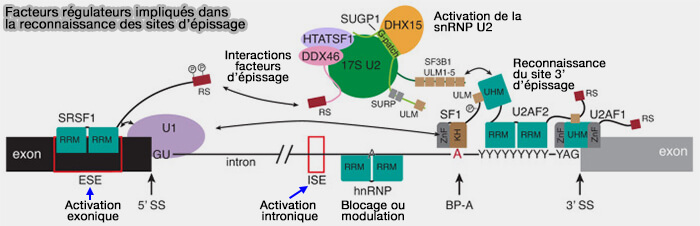
Facteurs régulateurs impliqués dans la reconnaissance des sites d’épissage
(Figure : vetopsy.fr d'après Tholen)
3. La di-snRNP U4/U6 et la snRNP U5 sont ensuite recrutées afin de former le spliceosome précatalytique.
La snARN U4 reste initialement appariée au snARN U6 et maintient celui-ci dans une conformation inactive empêchant une activation prématurée du centre catalytique.
La snARN U6 constitue l’un des principaux composants catalytiques du spliceosome.
Après dissociation de U4, le snARN U6 établit de nouveaux appariements avec le snARN U2 et avec le site 5′ d’épissage, conduisant à la formation du centre catalytique actif du spliceosome.
La snRNP U5 participe principalement à l’alignement des exons avant leur ligature.
Le snARN U5 interagit notamment avec les extrémités des exons et contribue à leur rapprochement lors de la seconde réaction de transestérification.
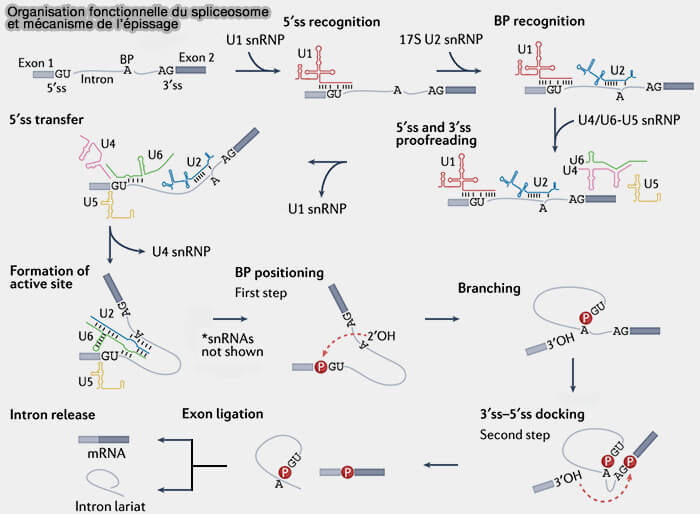
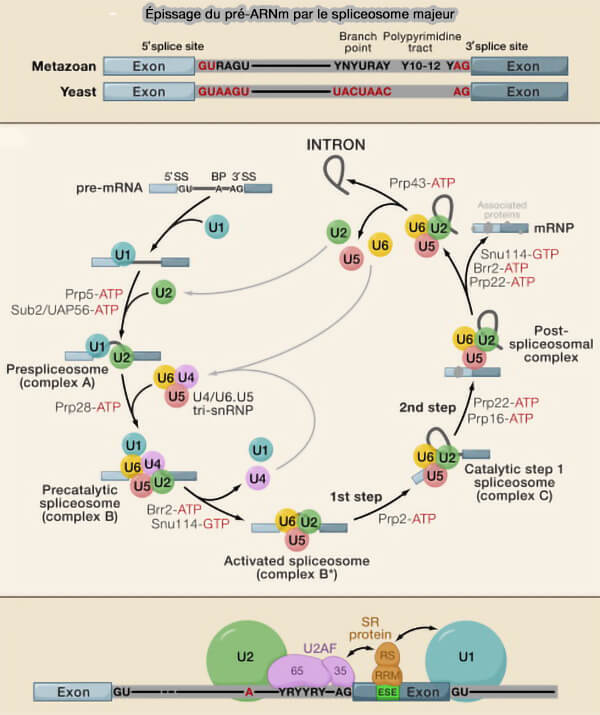
Organisation fonctionnelle du spliceosome et mécanisme de l’épissage
(Figure : vetopsy.fr adaptée d'après Rogalska et coll)
Une réaction de transestérification correspond à un échange de liaisons phosphodiester entre différents groupements hydroxyles d’une molécule d’ARN sans consommation directe d’ATP, contrairement à de nombreuses réactions enzymatiques de clivage ou de ligature des acides nucléiques.
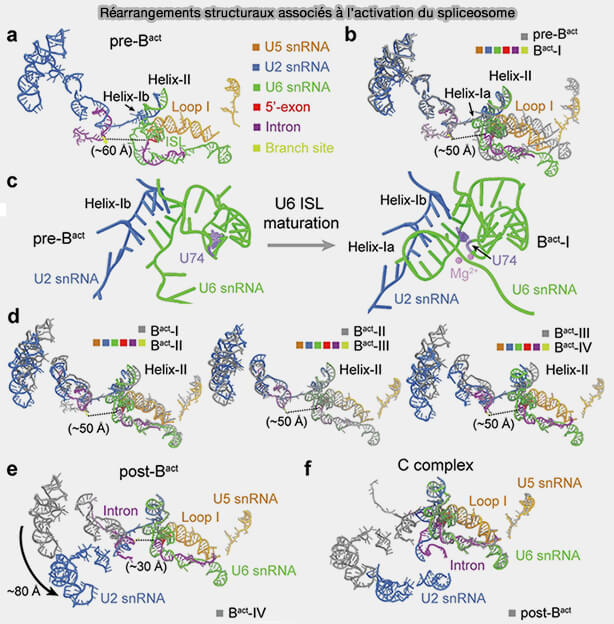
a. Lors de l’activation du spliceosome, le snARN U4 initialement apparié au snARN U6 est dissocié, permettant au snARN U6 d’établir de nouvelles interactions avec le snARN U2 et avec le site 5′ d’épissage.
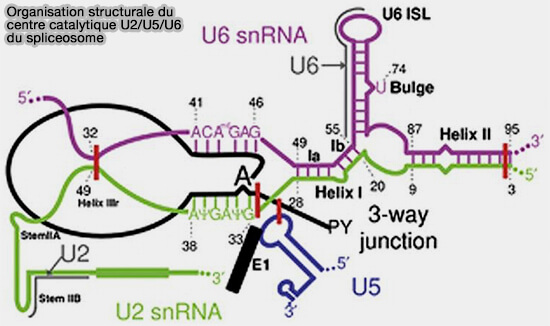
b. Ces réarrangements conduisent à la formation du cœur catalytique actif du spliceosome organisé autour des snARN U2 et U6.
Les interactions entre les snARN U2 et U6 génèrent plusieurs structures hélicoïdales conservées permettant l’organisation du site actif catalytique et la coordination de deux ions métalliques divalents, principalement Mg++, indispensables aux réactions de transestérification.
c. Cette organisation catalytique présente une forte parenté structurale et fonctionnelle avec les introns auto-épissants du groupe II, soutenant l’hypothèse d’une origine évolutive commune entre le spliceosome et ces ribozymes ancestraux.
Réarrangements structuraux associés à l’activation du spliceosome
(Figure : vetopsy.fr d'après Zhan et coll)
2. Lors de la première réaction de transestérification, les interactions entre les snARN U2, U6 et le pré-ARNm organisent le centre catalytique du spliceosome de manière à positionner correctement l’adénosine du point de branchement à proximité du site 5′ d’épissage.
Le groupement hydroxyle 2′OH porté par le ribose de cette adénosine attaque alors la liaison phosphodiester située à la jonction entre l’exon amont et l’intron au niveau du site donneur 5′ d’épissage.
Cette attaque nucléophile provoque le clivage de l’extrémité 5′ de l’intron et le transfert de cette extrémité sur l’adénosine du point de branchement.
L’intron forme alors une structure en lasso (lariat) dans laquelle son extrémité 5′ est reliée au groupement 2′OH de l’adénosine par une liaison phosphodiester 2′-5′ inhabituelle.
3. Lors de la seconde réaction de transestérification, l’exon amont libéré après la première réaction possède une extrémité 3′OH libre.
Ce groupement 3′OH attaque alors la liaison phosphodiester située au niveau du site accepteur 3′ d’épissage séparant l’intron de l’exon aval.
Cette seconde attaque nucléophile entraîne la ligature covalente des deux exons par une liaison phosphodiester classique 3′-5′ et provoque simultanément la libération de l’intron sous forme de lariat (lasso).
4. Bien que l’activité catalytique du spliceosome soit principalement portée par les snARN U2 et U6, de nombreuses protéines spliceosomales stabilisent l’organisation du centre catalytique et contrôlent les réarrangements structuraux successifs nécessaires à l’assemblage, à l’activation et au recyclage du spliceosome.
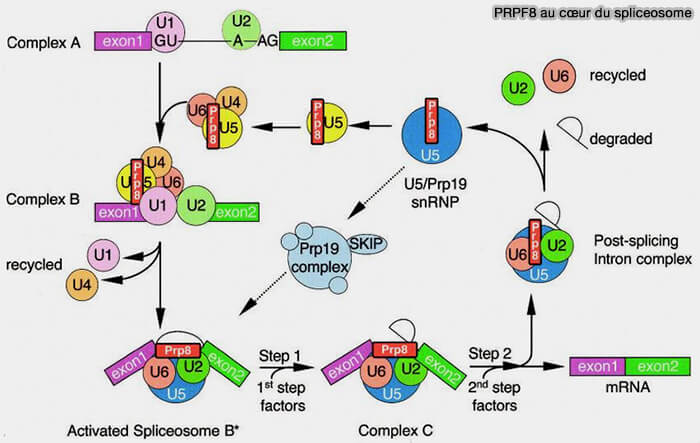
PRPF8 agit principalement comme une plateforme structurale organisant l’architecture du cœur actif du spliceosome et contribuant au positionnement correct des substrats ARN au cours des réactions de transestérification (Prp8 protein: At the heart of the spliceosome 2005)..
Elle participe également aux réarrangements conformationnels successifs du spliceosome permettant les transitions entre les différents états fonctionnels des complexes d’épissage.
PRPF8 au cceur du spliceosome
(Figure : vetopsy.fr d'après Grainger et Beggs)
Assemblage dynamique du spliceosome majeur
L’assemblage du spliceosome correspond à un processus hautement dynamique au cours duquel les différentes snRNP spliceosomales sont recrutées successivement sur le pré-ARNm puis réorganisées afin de former un centre catalytique actif capable de réaliser les réactions de transestérification.
Contrairement au ribosome qui constitue une structure relativement stable, le spliceosome est assemblé de manière transitoire sur chaque intron puis entièrement dissocié après l’épissage.
Ces enzymes utilisent l’énergie issue de l’hydrolyse de l'ATP pour remodeler dynamiquement les interactions ARN-ARN et ARN-protéines au cours du cycle d’épissage.
Parmi les principales hélicases du spliceosome figurent notamment Prp5/DDX46, Sub2/UAP56, Prp28/DDX23, Brr2/SNRNP200, Prp2/DHX16, Prp22/DHX8 et Prp43/DHX15, qui participent successivement :
à l’assemblage progressif des complexes spliceosomaux,
le complexe B activé (Bact), après dissociation des snRNP U1 et U4 et formation du centre catalytique actif organisé autour des snARN U2 et U6,
les complexes C catalytiques assurant les réactions de transestérification, avec le complexe C impliqué dans la première réaction et le complexe C* dans la seconde réaction,
les complexes post-catalytiques impliqués dans la dissociation et le recyclage du spliceosome.
b. Chaque transition structurale du spliceosome s’accompagne du recrutement et de la dissociation de nombreux facteurs protéiques spécialisés contrôlant l’assemblage, l’activation catalytique et le recyclage des complexes spliceosomaux.
3. Après la ligature des exons et la libération de l’intron sous forme de lariat, les différents composants du spliceosome sont dissociés puis recyclés afin de participer à de nouveaux cycles d’épissage.
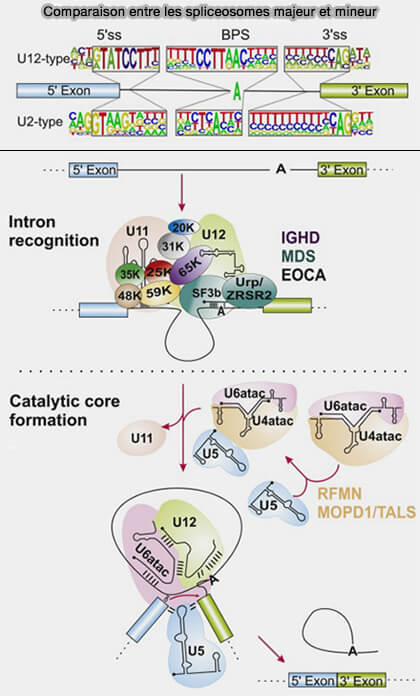
Spliceosome mineur
Chez certains eucaryotes, notamment chez les animaux, les plantes et plusieurs protistes, un spliceosome mineur distinct assure l’épissage d’une faible proportion d’introns appelés introns U12, représentant environ 0,3-0,5 % des introns chez l’humain (Minor spliceosome and disease 2018).
Structure du snARN U2
(Figure : vetopsy.fr d'après Verma et coll)
les voies de signalisation intracellulaire, notamment certaines protéines des voies MAPK, RAF ou protéines phosphatases et kinases régulatrices,
la maturation et l’épissage des ARN, notamment dans plusieurs composants du spliceosome mineur lui-même tels que RNPC3 (U11/U12-65K), ZRSR2 ou SNRNP48, contribuant à l’assemblage et au fonctionnement de cette machinerie d’épissage spécialisée.
Les facteurs d’épissage regroupent un ensemble de protéines et de complexes ribonucléoprotéiques régulateurs contrôlant la reconnaissance des sites d’épissage, l’assemblage du spliceosome et la sélection des exons au cours de la maturation des pré-ARNm.
Ils interviennent notamment dans la régulation de l’épissage alternatif qui permet de produire plusieurs isoformes d’ARNm à partir d’un même gène.




 centre catalytique).
centre catalytique).

{kind=link}
