


Acides nucléiques
ARN : types et fonctions
ARN non codants régulateurs de l’expression génique
1. Interférence par l’ARN (RNAi)
miRNA, siRNA et piRNA
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les miRNA, siRNA et piRNA sont des ARN non codants qui régulent l’expression génique par interférence avec les ARN cibles (RNAi), constituant une voie majeure de régulation post-transcriptionnelle.
ARN non codants régulateurs de l’expression génique
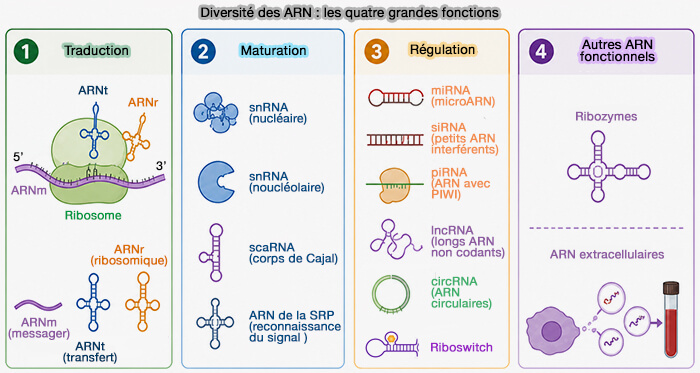
1. Les ARN non codants régulateurs de l’expression génique constituent une classe d’ARN qui modulent l’expression des gènes sans être traduits en protéines.
Ils agissent principalement en contrôlant la stabilité des ARN messagers, leur traduction ou l’état transcriptionnel de la chromatine.
(Figure : vetopsy.fr)
2. Les principales classes d’ARN non codants régulateurs sont les suivantes.
a. Les ARN impliqués dans l’interférence par l’ARN (RNAi) comprennent :
- les microARN (miRNA), qui inhibent la traduction ou favorisent la dégradation des ARNm.
- les petits ARN interférents (siRNA ou small interfering RNA), qui induisent la dégradation spécifique d’ARN cibles,
- les ARN associés aux protéines PIWI (piRNA ou piwi-interacting RNA), qui participent au contrôle des éléments transposables et à la protection du génome germinal.
b. Les autres ARN régulateurs, indépendants de l’interférence par l’ARN, comprennent :
- les ARN longs non codants (long non coding RNA), qui sont impliqués dans la régulation transcriptionnelle et l’organisation de la chromatine,
- les ARN circulaires (circRNA), qui participent à la régulation de l’expression génique, notamment en agissant comme éponges à microARN ou en modulant l’activité de protéines,
- les riboswitch, éléments structuraux d’ARN, généralement localisés dans les régions non traduites des ARNm, capables de lier directement des métabolites et de moduler l’expression génique par des changements de conformation.
ARN régulateurs impliqués dans l’interférence par l’ARN (RNAi)
Les ARN régulateurs impliqués dans l’interférence par l’ARN (RNA interference, RNAi) constituent une catégorie de petits ARN non codants qui contrôlent l’expression génique par des mécanismes de silençage post-transcriptionnel.

Le silençage post-transcriptionnel correspond à l’ensemble des mécanismes qui régulent l’expression des gènes en agissant sur les ARN messagers (ARNm) après leur transcription, notamment en inhibant leur traduction ou en favorisant leur dégradation.
1. L’interférence par l’ARN constitue une forme particulière de ce silençage, reposant sur l’action de petits ARN guides chargés sur une protéine de la famille Argonaute ou PIWI, qui reconnaît un ARN cible par complémentarité de séquence et dirige :
- soit son clivage,
- soit l’inhibition de sa traduction.
Outre l’interférence par l’ARN (RNAi), plusieurs autres mécanismes de silençage post-transcriptionnel peuvent réguler l’expression des gènes en agissant sur les ARN messagers ( dégradation et surveillance des ARN).
dégradation et surveillance des ARN).
2. Les RNAi comprennent notamment :
- les microARN (miRNA), qui répriment l’expression de nombreux ARNm par appariement partiel,
- les petits ARN interférents (siRNA), souvent issus d’ARN double brin et impliqués dans l’interférence par l’ARN,
- les ARN associés aux PIWIi (piRNA), associés aux protéines PIWI et spécialisés dans le contrôle des transposons, notamment dans la lignée germinale.
3. Ces petits ARN régulateurs agissent au sein de complexes effecteurs qui utilisent l’appariement de bases pour reconnaître des ARN cibles et provoquer leur dégradation ou leur inhibition traductionnelle.
microARN (miRNA)
Rôles de miRNA
Les microARN (miRNA) sont de petits ARN non codants d’environ 21-23 nucléotides qui régulent l’expression génique au niveau post-transcriptionnel.
Ils agissent principalement en inhibant la traduction des ARN messagers ou en favorisant leur dégradation après reconnaissance de séquences complémentaires présentes dans les ARNm cibles.

Les miRNA sont répertoriés dans : miRBase: the microRNA database.
Les miRNA constituent ainsi un réseau majeur de régulation post-transcriptionnelle, chaque miRNA pouvant contrôler simultanément l’expression de nombreux ARN messagers différents et participer à la régulation coordonnée de voies biologiques complexes telles que :
- le développement, comme miR-430 qui régule la transition maternelle-zygotique chez le poisson zèbre en éliminant les ARNm maternels,
- la prolifération cellulaire, comme miR-21 qui cible la phosphatase PTEN et favorise l’activation de la voie PI3K/Akt,
- la différenciation cellulaire, comme miR-1 et miR-133 qui régulent la différenciation musculaire en ciblant SRF (Serum response factor), HDAC4 et d’autres facteurs myogéniques,
- l’homéostasie cellulaire, comme miR-34, régulé par p53, qui contrôle l’apoptose en ciblant BCL2 et l’arrêt du cycle cellulaire en ciblant notamment CDK6 et des régulateurs du point de contrôle G1/S.
Biogenèse et mécanisme d’action des miRNA
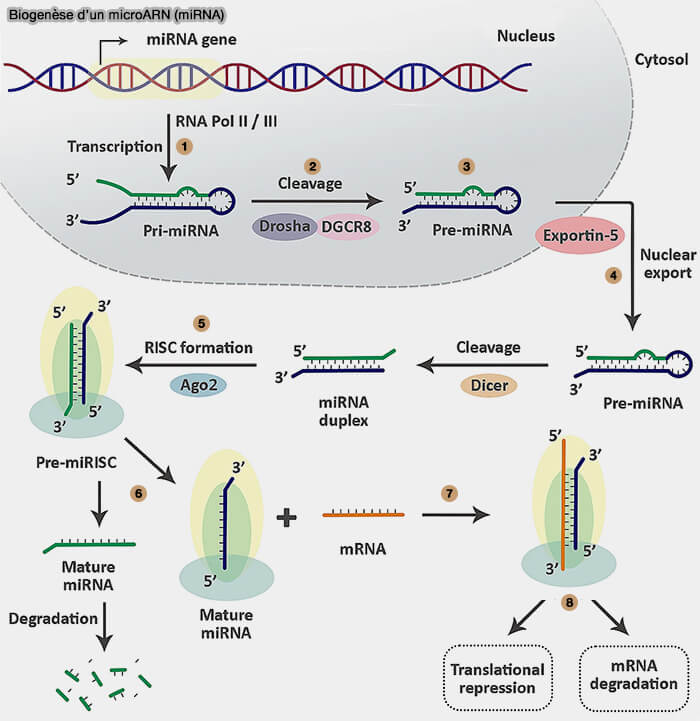
Les miRNA sont générés par une succession d’étapes de maturation enzymatique conduisant à leur incorporation dans le complexe RISC, qui assure ensuite la reconnaissance spécifique des ARN messagers cibles et la régulation de leur expression.
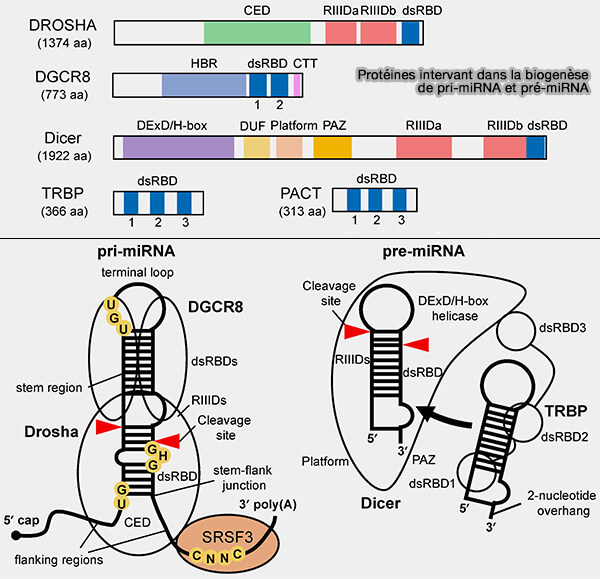
1. Les miRNA sont transcrits par l’ARN polymérase II sous forme de longs transcrits appelés pri-miRNA (primary microRNA) de plusieurs centaines à milliers de nucléotides, contenant une structure caractéristique en tige-boucle qui n’est pas toujours parfaitement appariée (mésappariements, bulges et petites irrégularités structurales).
2. Dans le noyau, ces transcrits sont reconnus par le complexe microprocesseur constitué de l’endoribonucléase Drosha et de la protéine DGCR8.
- DGCR8 reconnaît la structure en tige-boucle du pri-miRNA et se fixe sur les régions double brin de la tige et positionne correctement l’endoribonucléase Drosha.
- Drosha, RNase III, clive le pri-miRNA pour générer un pré-miRNA (pre-miRNA) d’environ 60-70 nucléotides conservant la structure en épingle à cheveux (Sperm epigenetics and male infertility: unraveling the molecular puzzle 2024).
Le complexe Drosha-DGCR8 clive les deux brins de la tige du pri-miRNA à des positions légèrement décalées, ce qui génère un pré-miRNA dont chaque extrémité du duplex possède deux nucléotides non appariés du côté 3′ (overhang ou surplomb).
(Figure : vetopsy.fr d'après encyclopedia.pub)
3. Le pré-miRNA est ensuite exporté vers le cytoplasme par la protéine Exportin-5 en association avec Ran-GTP.
4. Dans le cytoplasme, l’endoribonucléase Dicer, qui est aussi une RNASE III, reconnaît le surplomb de deux nucléotides à l’extrémité 3′ du pré-miRNA et clive la tige-boucle à une distance définie de cette extrémité, générant un duplex d’ARN d’environ 22 nucléotides composé de deux brins complémentaires, miRNA et miRNA* avec deux bases non appariées du côté 3'.
a. Après clivage du pré-miRNA par Dicer, les deux brins du duplex généré proviennent respectivement des bras 5′ et 3′ de la structure en tige-boucle.
- Les microARN matures sont ainsi désignés par les suffixes 5p et 3p, correspondant à leur origine structurale dans le pré-miRNA, par exemple miR-21-5p et miR-21-3p.
- L’un des deux brins est généralement chargé préférentiellement dans le complexe RISC pour former le brin guide fonctionnel, tandis que l’autre correspond au brin passager.
(Figure : vetopsy.fr d'après Lonugett)
{kind=link}
b. Cependant, les deux formes peuvent parfois être stables, associées à Argonaute et réguler des ARN messager (ARNm) différents, ce qui explique l’abandon progressif de l’ancienne nomenclature utilisant le symbole *.
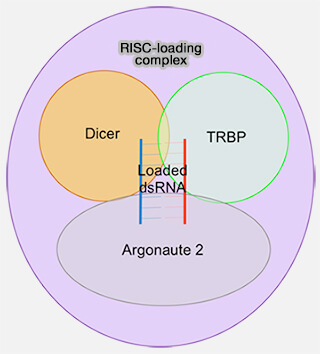
5. Ce duplex est pris en charge par le complexe RLC (RISC-loading complex), un complexe de chargement comprenant notamment :
- Dicer,
- TRBP ou PACT, cofacteurs alternatifs de Dicer qui se lient au même site ou à des sites proches et sont mutuellement exclusifs au sein d’un même complexe, modulant différemment l’activité de Dicer et l’efficacité du chargement sur une protéine Argonaute (AGO1-4), également recrutée au sein de ce complexe.
a. Le chargement du brin guide dans Argonaute au sein de ce complexe conduit à la formation du complexe RISC (RNA-induced silencing complex), qui correspond à la forme active minimale constituée d’Argonaute associée au miRNA guide, tandis que Dicer, TRBP et PACT n’interviennent que de manière transitoire lors du chargement et ne font pas partie du RISC mature.
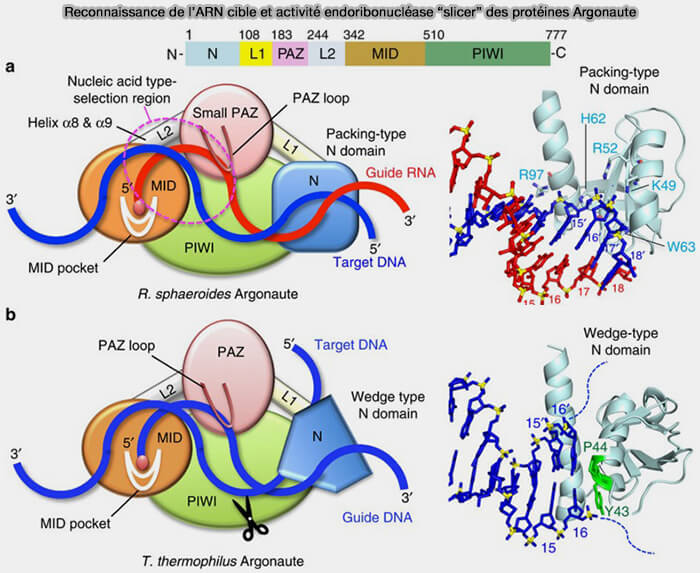
b. La protéine Argonaute, notamment Ago2, grâce à son domaine PAZ (Drosophila Piwi, Arabidopsis Argonaute-1, and Arabidopsis Zwille), reconnaît le surplomb, pour former un complexe pré-miRISC dans lequel les deux brins du duplex sont encore présents.
(Figure : vetopsy.fr d'après Hosseini et coll)
6. Le complexe pré-miRISC (preRNA-induced silencing complex) est maturé par sélection d’un brin guide (Structural basis for the recognition of guide RNA and target DNA heteroduplex by Argonaute 2016).
a. Le brin guide (miRNA) reste associé à Argonaute dans le complexe miRISC mature.
b. L’autre brin, appelé brin passager (miRNA*), est généralement dégradé et cette élimination concerne le duplex miRNA/miRNA* produit par Dicer, avant toute reconnaissance d’un ARNm cible.
Le duplex miRNA/miRNA* est initialement maintenu par de multiples appariements de bases sur toute sa longueur et deux cas peuvent se produire.
(Figure : vetopsy.fr d'après Miyoshi et coll)
b1. Lorsque les deux brins du duplex sont fortement appariés, Ago2 peut cliver le brin passager par l'activité dite " slicer ", portée par le domaine PIWI d’Ago2, qui adopte le même repliement structural et le même principe catalytique (coordination d’ions Mg++ et clivage du brin cible d’un duplex) que les endoribonucléases de type RNase H.
L’activité endoribonucléase dite " slicer " correspond à la capacité spécifique de certaines protéines Argonaute (notamment Ago2) à cliver un ARN cible en utilisant un petit ARN guide (miRNA ou siRNA) comme matrice d’appariement, ce qui la distingue des autres endonucléases qui reconnaissent directement des séquences ou des structures sans intermédiaire guide.
- Elle introduit une coupure unique au centre du brin passager, en face des nucléotides 10-11 du brin guide, ce qui interrompt la continuité de son squelette phosphodiester qui n’est alors plus une chaîne unique mais deux fragments indépendants, incapables de maintenir les contraintes mécaniques nécessaires à un appariement stable avec le brin guide.
- Cette rupture centrale entraîne une perte de stabilité du duplex, provoque son désembobinage (unwinding) et permet la dissociation complète des deux brins, suivie de la dégradation rapide des fragments du brin passager par des exoribonucléases.
b2. Lorsque le duplex contient des mésappariements ou des irrégularités structurales, cas le plus fréquent chez les miRNA, le brin passager est plutôt dissocié par déstabilisation thermodynamique sans clivage par l’activité " slicer " d’Ago2, puis rapidement dégradé par les exoribonucléases cytoplasmiques précédentes.
7. Le miRNA guide le complexe RISC vers les ARN messagers (ARNm) cibles grâce à un appariement avec des séquences complémentaires, le plus souvent localisées dans la région 3′UTR.
a. La spécificité de cette reconnaissance repose principalement sur une courte région du miRNA appelée séquence seed (nucléotides 2-8), essentielle pour la spécificité de la reconnaissance.
8. Selon le degré de complémentarité entre le miRNA et l’ARN cible, deux mécanismes principaux peuvent être observés, cette différence reposant sur la capacité du duplex miRNA-ARNm à positionner correctement la région centrale du miRNA (nucléotides 10-11) dans le site catalytique du domaine PIWI d’Argonaute
8a. Lorsque l’appariement est partiel, le complexe RISC inhibe la traduction de l’ARNm et favorise sa déstabilisation progressive, par recrutement de protéines effectrices de type GW182/TNRC6 associées à Argonaute.
a. Ces protéines recrutent notamment le complexe CCR4-NOT, responsable de la déadénylation de la queue poly(A), ce qui diminue la stabilité de l’ARNm et favorise son retrait des polysomes.
b. Après déadénylation, plusieurs devenirs sont possibles.
- L’ARNm peut subir un décapping par les complexes DCP1/DCP2, puis être dégradé dans le sens 5′ → 3′ par l’exoribonucléase XRN1.
- L’ARNm peut également être dirigé vers les P-bodies, qui concentrent des facteurs de répression, de décapping et de dégradation, et où il peut être soit stocké transitoirement, soit engagé dans une dégradation cytoplasmique.
- Lors de certains stress cellulaires, une partie des ARNm réprimés peut aussi être redistribuée vers des granules de stress, qui correspondent plutôt à des sites transitoires de stockage et de tri des ARNm non traduits qu’à des sites principaux de dégradation
8b. Lorsque la complémentarité est presque parfaite, la protéine Argonaute peut cliver directement l’ARNm cible en face des nucléotides 10-11 du miRNA guide grâce à son activité endonucléase " slicer ", portée par le domaine PIWI.
Ce clivage génère deux fragments d’ARN rapidement pris en charge par les systèmes cytoplasmiques de dégradation, notamment par des exoribonucléases agissant dans les sens 5′ → 3′ et 3′ → 5′.
Remarque : la protéine Argonaute intervient donc à deux niveaux distincts dans le processus :
- en amont, lors du chargement du duplex miRNA/miRNA*, où elle peut cliver le brin passager via son activité " slicer " afin de permettre la formation d’un complexe actif,
- en aval, au sein du RISC mature, où elle assure la reconnaissance des ARNm cibles et, selon le degré de complémentarité, leur répression ou leur clivage.
Petits ARN interférents (siRNA ou small interfering RNA)
Les petits ARN interférents ou siRNA (small interfering RNA) sont de courts ARN double brin d’environ 21-23 nucléotides qui conduisent au silençage spécifique de gènes par dégradation d’ARN cibles.
1. Les siRNA proviennent généralement du clivage d’ARN double brin longs (dsRNA) qui peuvent être d’origine :
- virale lors d’infections produisant des ARN double brin,
- endogène, notamment à partir de séquences répétées inversées générant des structures en tige-boucle, de transcrits issus de transposons pouvant s’apparier entre eux, ou encore par hybridation entre ARN sens et antisens produisant un duplex d’ARN,
- expérimentale, dans les approches d’interférence par ARN utilisées en biologie moléculaire.
2. Comme pour les microARN, la biogenèse et le mécanisme d’action des siRNA reposent sur leur chargement dans le complexe RISC, qui dirige le siRNA guide vers les ARN cibles par appariement complémentaire.
- Contrairement aux miRNA, cette complémentarité est généralement quasi parfaite, ce qui permet à la protéine Argonaute-2 (Ago2) de cliver directement l’ARN cible en face des nucléotides 10-11 du siRNA guide grâce à son activité endonucléase dite " slicer ".
- L’ARN clivé est ensuite rapidement dégradé par les ribonucléases cellulaires, ce qui conduit à une diminution efficace du niveau de l’ARN cible et donc à un silençage post-transcriptionnel du gène correspondant.
ARN associés aux PIWI (piARN ou piwi-interacting RNA)
Les piRNA (piwi-interacting RNA) sont une classe de petits ARN non codants d’environ 24-31 nucléotides qui s’associent aux protéines de la sous-famille PIWI de la famille Argonaute et jouent un rôle majeur dans la répression des éléments transposables, en particulier dans les cellules germinales.
- La biogenèse des piRNA a été principalement caractérisée chez la drosophile, où les différents cofacteurs de maturation, de chargement et d’amplification ont été identifiés, notamment dans le mécanisme de ping-pong impliquant les protéines PIWI Aubergine et Ago3.
- Chez les vertébrés, les principes fondamentaux de la voie piRNA sont conservés, bien que les protéines impliquées correspondent à des homologues spécifiques tels que MILI, MIWI et MIWI2 chez les mammifères.
Vous pouvez lire : Silencing of Transposable Elements by piRNAs in Drosophila: An Evolutionary Perspective (2017).
Biogenèse des piRNA
1. Contrairement aux miRNA et aux siRNA, les piRNA dérivent de longs transcrits simples brins issus de régions génomiques appelées clusters piRNA, riches en fragments de rétrotransposons.
Ces régions sont transcrites pour produire de longs précurseurs d’ARN qui servent de source aux piRNA, et leur maturation est indépendante de l’endonucléase Dicer, qui agit spécifiquement sur des ARN double brin.
2. Ces précurseurs piRNA sont exportés du noyau vers le cytoplasme par une machinerie d’export des ARN, impliquant notamment le complexe NXF1/NXT1, principal récepteur d’export des ARN messager (ARNm) et de certains longs ARN ribonucléoprotéiques (lncRNA).
La différence importante est que NXF1/NXT1 ne fonctionne pas comme la karyophérine Exportin-5 en dépendante de Ran-GTP, mais utilise une voie distincte d’export reposant principalement sur des interactions directes avec les nucléoporines FG du pore nucléaire.
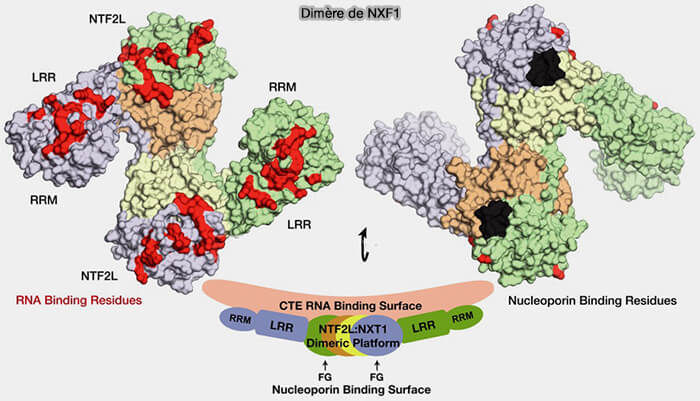
(Figure : vetopsy.fr d'après Aibara et coll)
3. Dans le cytoplasme, les longs précurseurs sont clivés en fragments courts par une endonucléase appelée Zucchini (Zuc), appelée PLD6 chez les vertébrés, enzyme associée à la membrane externe des mitochondries (OMM) appartenant à la famille PLD (phospholipase D), qui génère les extrémités 5′ des futurs piRNA (Zucchini: the key ingredient to unveil piRNA precursor processing 2020).
- Dans les cellules germinales de la drosophile, ces transcrits sont ensuite dirigés vers des granules cytoplasmiques spécialisés appelés Yb bodies, où se concentrent plusieurs facteurs de la biogenèse primaire des piRNA, notamment les protéines Armitage (Armi), Vreteno (Vret), Yb et la protéine Papi (Partner of PIWI), qui facilitent le transfert, le chargement et la stabilisation des fragments pré-piRNA sur les protéines PIWI en cours de maturation (Structural insights into the sequence-specific recognition of Piwi by Drosophila Papi 2018).
- Chez les vertébrés, les protéines de la voie piRNA se concentrent dans des granules cytoplasmiques périmitochondriaux appelés nuage (ou germ granules), structures ribonucléoprotéiques spécialisées où s’organisent une partie de la maturation, du chargement et de l’amplification des piRNA.
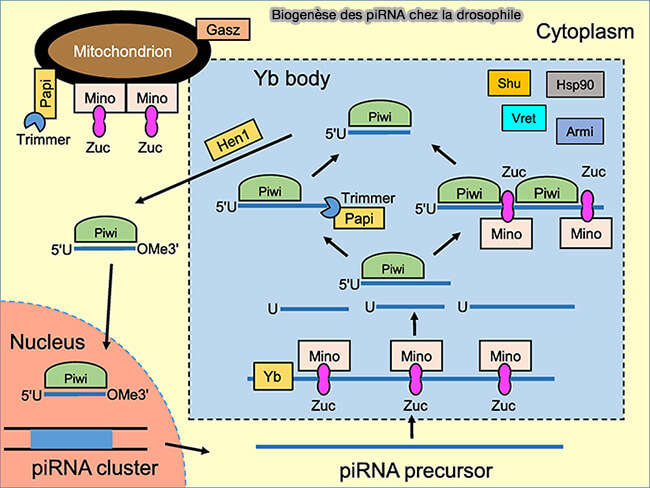
(Figure : vetopsy.fr d'après Luo et Lu)
4. Ces fragments courts sont ensuite chargés sur des protéines de la sous-famille PIWI des Argonautes, telles que PIWIL1 (MIWI), PIWIL2 (MILI) ou PIWIL4 (MIWI2) chez les vertébrés, pour former des complexes piRNA-PIWI capables de reconnaître des ARN dérivés de rétrotransposons par complémentarité de séquence.
a. Le chargement et la stabilisation de ces complexes nécessitent l’intervention de chaperons moléculaires, notamment Shutdown (Shu) et Hsp90, qui facilitent les changements conformationnels des protéines PIWI et l’incorporation correcte des fragments pré-piRNA dans leur poche de liaison.
b. Une fois liés à PIWI, l’extrémité 3′ est :
- raccourcie (trimming) par des exonucléases 3′ ➞ 5′ appelées trimérases, notamment Trimmer/Nibbler chez la drosophile et PNLDC1 (Poly(A)-specific ribonuclease-like domain-containing protein 1) chez les vertébrés, qui ajustent précisément la longueur de l’extrémité 3′ des piRNA en maturation,
- puis méthylée en 2′-O par la méthyltransférase HENMT1 (homologue de Hen1), ce qui protège les piRNA contre les exoribonucléases et stabilise leur accumulation dans les complexes PIWI.
5. Plusieurs protéines supplémentaires participent à l’organisation de la voie piRNA dans les cellules germinales et apparaissent fréquemment dans la biogenèse des piRNA (The RNA helicase MOV10L1 binds piRNA precursors to initiate piRNA processing 2015).
- Chez les vertébrés, l’hélicase à ARN MOV10L1 joue un rôle central dans l’initiation de la biogenèse primaire en remodelant les longs précurseurs d’ARN issus des clusters piRNA et en facilitant leur transfert vers l’endonucléase mitochondriale PLD6/Zucchini.
- La protéine Gasz, localisée à la membrane externe des mitochondries, contribue quant à elle à l’organisation du compartiment périmitochondrial où se déroule la maturation des piRNA, en servant de plateforme d’assemblage pour plusieurs facteurs de la voie piRNA.
- Enfin, plusieurs protéines à domaine Tudor (TDRD), telles que TDRD1, TDRD9 ou TDRD12, participent à l’organisation des complexes ribonucléoprotéiques piRNA dans les granules germinaux et reconnaissent les protéines PIWI modifiées, contribuant ainsi à la stabilisation des complexes piRNA et au fonctionnement du cycle d’amplification.
6. Les complexes piRNA-PIWI agissent au niveau post-transcriptionnel, en clivant directement les ARN de transposons dans le cytoplasme, dans le cytoplasme grâce à l’activité " slicer " des protéines PIWI, mais aussi au niveau transcriptionnel.
- Une fraction des complexes piRNA-PIWI est transportée vers le noyau grâce à un signal de localisation nucléaire (NLS) porté par certaines protéines PIWI et reconnu par la machinerie d’import nucléaire dépendante des importines.
- Une fois dans le noyau, ces complexes peuvent reconnaître les transcrits naissants de transposons par complémentarité de séquence et recruter des complexes épigénétiques responsables de la méthylation de l’ADN et de modifications répressives de la chromatine, notamment la triméthylation de H3K9, conduisant à la la formation d’hétérochromatine et à la répression transcriptionnelle durable de ces éléments mobiles.
Cycle d'amplification dans les cellules germinales
Dans les cellules germinales, les piRNA issus de la biogenèse primaire peuvent initier deux mécanismes de production secondaire :
- le cycle d’amplification " ping-pong ", fondé sur des clivages réciproques entre protéines PIWI,
- une voie alternative dépendante de l’endonucléase Zucchini, appelée phasing, qui produit de manière séquentielle plusieurs piRNA à partir d’un même transcrit précurseur (One Loop to Rule Them All: The Ping-Pong Cycle and piRNA-Guided Silencing 2016).
Dans les cellules germinales, le cycle " ping-pong " repose sur l’activité endonucléolytique de protéines PIWI complémentaires, Aubergine et Ago3 chez la drosophile (respectivement MILI et MIWI2 chez les vertébrés), qui génèrent de nouveaux piRNA par clivages réciproques de transcrits de transposons (One Loop to Rule Them All: The Ping-Pong Cycle and piRNA-Guided Silencing 2016).
1. Le cycle " ping-pong " est un mécanisme d’amplification fondé sur l’activité endonucléolytique des protéines PIWI.
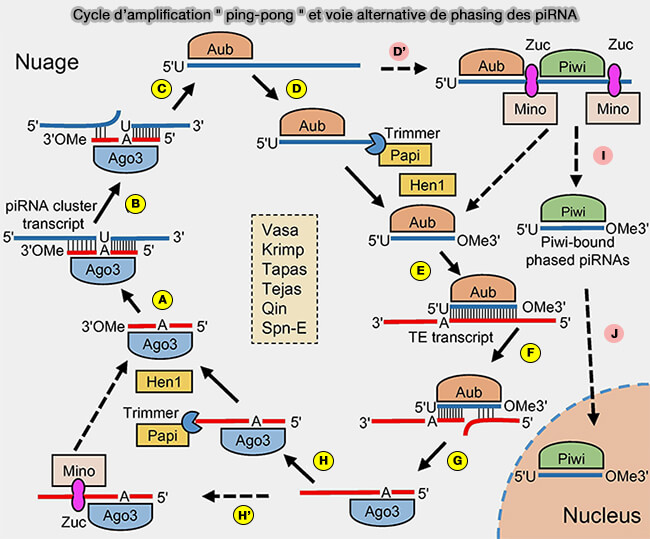
(Figure : vetopsy.fr d'après Luo et Lu)
a. Il implique une succession de clivages réciproques entre les protéines Ago3 et Aubergine (Aub), (respectivement MIWI2 et MILI chez les vertébrés), chacune utilisant les piRNA qu’elle porte pour reconnaître et cliver des ARN complémentaires.
- A. Un piRNA sens, i.e. qui possède une séquence identique (ou très proche) à celle du transcrit de transposon, lié à Ago3, reconnaît par complémentarité un transcrit antisens issu d’un cluster piRNA.
- B. Le transcrit du cluster piRNA est clivé par Ago3, générant l’extrémité 5′ d’un nouveau piRNA antisens généralement caractérisé par un uracile en position 1 (1U bias)
- C. L’intermédiaire piRNA antisens est ensuite chargé sur Aubergine (Aub).
- D. L’extrémité 3′ du piRNA antisens est raccourcie à sa longueur mature par l’exonucléase Trimmer, avec l’aide de son cofacteur Papi.
D′. Une voie alternative ou parallèle peut impliquer un clivage par Zucchini ( voie alternative ou phasing).
- E. Après méthylation en 2′-O par Hen1, le piRNA antisens mature lié à Aub reconnaît un transcrit de transposon.
- F. Le transcrit de transposon est clivé par Aub, générant l’extrémité 5′ d’un nouveau piRNA sens souvent caractérisé par une adénine en position 10 (10A bias), signature du cycle ping-pong.
- G. L’intermédiaire piRNA sens est ensuite chargé sur Ago3.
- H. Son extrémité 3′ est raccourcie par Trimmer, avec le cofacteur Papi.
- H′. Alternativement, l’intermédiaire sens peut être clivé par Zucchini (Zuc).
Cette voie peut produire un piRNA sens mature lié à Ago3, qui réamorce le cycle en revenant à l’étape A, mais aussi générer des piRNA phasés associés à Piwi, qui suivent ensuite la branche I-J vers la maturation et l’import nucléaire.
b. Ce mécanisme produit ainsi de nouveaux piRNA sens et antisens à partir des fragments générés par ces clivages.
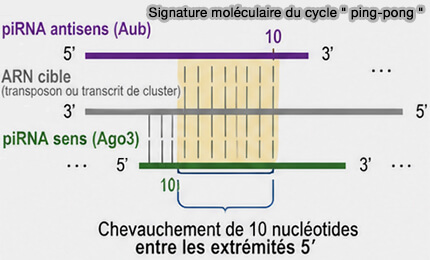
(Figure : vetopsy.fr)
- Les extrémités 5′ des piRNA produits sont décalées de 10 nucléotides, car les protéines PIWI clivent l’ARN cible en face du dixième nucléotide du piRNA guide.
- Le fragment généré devient alors le point de départ du piRNA suivant, ce qui crée un chevauchement caractéristique de 10 nucléotides entre piRNA complémentaires, signature moléculaire du cycle " ping-pong " .
- Cette amplification permet d’augmenter rapidement la quantité de piRNA dirigés contre les transcrits de transposons, renforçant ainsi leur élimination et contribuant à la protection du génome des cellules germinales.
2. Une voie alternative (phasing) peut également intervenir lors de la maturation des piRNA, mais repose sur un principe différent du cycle ping-pong.
Au lieu de générer un seul piRNA par clivage ciblé, un précurseur d’ARN est clivé successivement le long du même transcrit, produisant une série de piRNA contigus.
- Chaque nouveau clivage définit l’extrémité 5′ du piRNA suivant, ce qui entraîne la formation d’une cascade de piRNA régulièrement espacés le long du transcrit précurseur.
- Cette production séquentielle permet de générer rapidement plusieurs piRNA à partir d’un même ARN et peut alimenter à la fois la voie cytoplasmique de ping-pong et la voie nucléaire associée à Piwi
D'. La bifurcation commence à l’étape D′ car certains intermédiaires d’ARN, au lieu d’être uniquement maturés par trimming, sont clivés de manière séquentielle par Zucchini (Zuc), ce qui peut générer soit des piRNA réinjectés dans le cycle ping-pong, soit des piRNA phasés associés à Piwi.
La bifurcation commence à D' car, au lieu d’être raccourcis par l’exonucléase Trimmer, certains intermédiaires d’ARN sont clivés par l’endonucléase mitochondriale Zucchini (Zuc), ce qui génère :
- soit des piRNA liés à Aub, qui peuvent réintégrer la voie canonique du cycle " ping-pong " ,
- soit des piRNA phasés associés à Piwi.
I. Ces piRNA sont maturés par trimming puis méthylation en 2′-O par Hen1, ce qui stabilise leur extrémité 3′.
J. Les piRNA liés à Piwi sont ensuite importés dans le noyau, où ils participent à la répression transcriptionnelle des loci de transposons.
3. Les protéines situées au centre de la figure, notamment Vasa, Krimper (Krimp), Tapas, Tejas, Qin et Spindle-E (Spn-E), organisent et stabilisent le cycle " ping-pong " dans les granules germinaux en coordonnant le recrutement, l’assemblage, le remodelage et la stabilisation des complexes piRNA-PIWI
Le tableau suivant résume les principales caractéristiques des petits ARN impliqués dans les mécanismes d’interférence par l’ARN.
| Caractéristique | MicroARN (miRNA) |
Petits ARN interférents (siRNA) |
ARN associés aux PIWI (piRNA) |
|---|---|---|---|
| Taille | ~21-23 nt | ~21-23 nt | ~24-31 nt |
| Origine | Transcrits endogènes formant des structures en tige-boucle (pri-miRNA) |
ARN double brin longs (dsRNA)
|
Transcrits longs issus de régions génomiques riches en transposons |
| Maturation | Drosha → Exportin-5 → Dicer |
Dicer | Indépendante de Dicer |
| Complexe effecteur | miRISC (Argonaute) | RISC (Argonaute) | Complexes piRNA-PIWI |
| Appariement avec la cible |
Généralement partiel |
Généralement quasi parfait |
variable |
| Effet principal | Inhibition de la traduction et déstabilisation des ARNm |
Cllivage direct de l’ARN cible |
Répression des éléments transposables |
| Localisation principale |
Cytoplasme | Cytoplasme | Cytoplasme et noyau des cellules germinales |
ARN régulateurs indépendants de l’interférence par l’ARN
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation