


Nucléotides : structure
Bases azotées
: propriétés
- Biochimie
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les bases azotées possèdent des propriétés chimiques et structurales spécifiques qui déterminent leur réactivité, leur tautomérie et leur capacité à former des liaisons hydrogène.
Les nucléotides contiennent une base azotée hétérocyclique, appelée nucléobase, qui détermine en grande partie leurs propriétés chimiques et leur capacité de reconnaissance moléculaire.
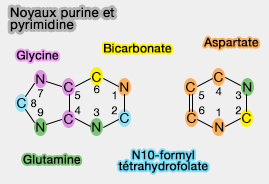
(Figure : vetopsy.fr)
Les nucléobases biologiques appartiennent à deux grandes familles structurales :
- les bases puriques, dérivées du noyau purine,
- les base pyrimidiques, dérivées du noyau pyrimidine.
Dans les acides nucléiques, ces bases correspondent principalement à :
- l'adénine (A) ou la guanine (G), pour les bases puriques,
- la cytosine (C), la thymine (T) spécifique à l'ADN, ou l'uracile (U), spécifique à l'ARN pour les soit une bases pyrimidiques.
Propriétés des bases puriques et pyrimidiques
Par leus propriétés chimiques, les bases azotées déterminent :
- les règles d’appariement, mais aussi la stabilité, la conformation et la capacité des acides nucléiques à stocker et transmettre l’information génétique,
- des liaisons spécifiques qui jouent également un rôle clé dans des processus biologiques tels que la catalyse enzymatique, la signalisation cellulaire et le métabolisme énergétique.
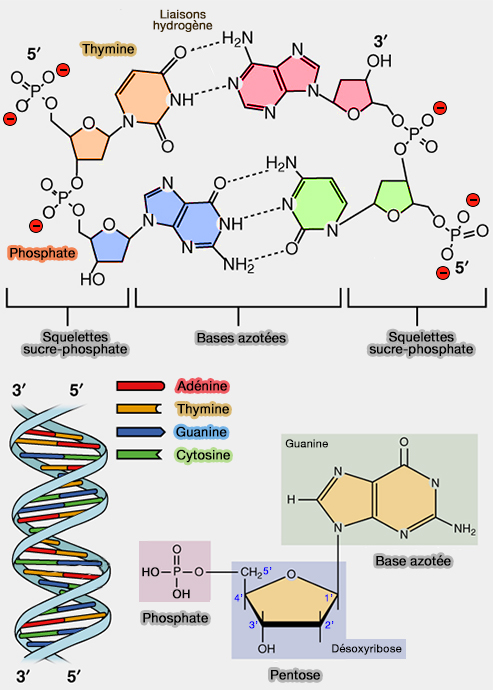
(Figure : vetopsy.fr d'après OpenStax)
{kind=link}
Noyau aromatique
Le caractère aromatique du noyau des bases azotées leur confère plusieurs propriétés essentielles.
1. La stabilité structurale tridimentionnelle des acides nucléiques repose sur :
- des interactions π–π entre bases empilées, ce qui influence la conformation et la fonction des brins d’ADN ou d’ARN.
- la résistance à l'oxydation, liée à la délocalisation des électrons π dans le système aromatique, contribuant à la durabilité des acides nucléiques dans les cellules.
2. Le rôle dans la reconnaissance moléculaire est assuré, en particulier, par des des liaisons hydrogène spécifiques.
3. La capacité à subir des réactions de substitution, comme la désamination de la cytosine en uracile, est un facteur de mutation qui justifie l’existence de mécanismes cellulaires de réparation de l’ADN.
Remarque : l'absorption caractéristique dans l'ultraviolet, surtout à 260 nm, qui varie avec le pH du miliieu, est due au système de doubles liaisons conjuguées, ce qui permet leur identification et leur dosage.
Tautomères
Les tautomères sont des formes chimiques alternatives d’une même molécule résultant du déplacement d’un proton et d’une réorganisation des doubles liaisons.
Dans les bases azotées, ce phénomène est favorisé par la présence de de groupes hydroxyles ($\ce{-OH}$) ou amines ($\ce{-NH2}$) et correspond le plus souvent à une tautomérie amide/acide imidique, forme particulière de tautomérie céto-énol.
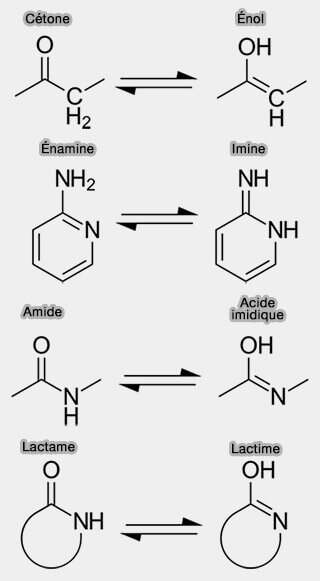
(Figure : vetopsy.fr)
$\ce{R−C(=O)−NH2}$ $\leftrightharpoons$ $\ce{R−C(OH)=NH}$.

La tautomérie, cas particulier de l'isomérie de fonction, est étudiée dans un chapitre spécifique.
1. Les tautomères sont très rares (≈ 10-4 à 10-5 des bases), mais ils peuvent influencer plusieurs propriétés biologiques importantes :
- l’appariement des bases,
- la catalyse enzymatique,
- la reconnaissance moléculaire.
2. Dans certains ribozymes, des formes tautomères peuvent contribuer à la stabilisation de l’état de transition des réactions catalytiques.
Pyrimidine
Les formes tautomères possibles pour les bases pyrimidiques sont :
- la forme lactame, $\ce{R−C(=O)−NH2}$, majoritaire et stable au pH physiologique,
- la forme lactime, $\ce{R−C(OH)=NH}$, minoritaire mais biologiquement significative,
Purines
1. Les bases puriques présentent également plusieurs formes tautomères :
- la forme lactame, $\ce{R−C(=O)−NH2}$, prédominante au pH physiologique,
- les formes imine, imino ($\ce{=NH)$) ou énol ($\ce{C–OH}$), plus rares et dépendent de la position du proton mobile.
Les formes imines des purines existent en très faibles proportions, mais elles ont une importance biologique non négligeable.
2. En effet, ces formes rares des purines, bien qu’instables, peuvent avoir des conséquences importantes.
a. Les tautomères modifient temporairement les propriétés d'appariement des bases, notamment leur capacité à former des liaisons hydrogène.
- Une base sous forme tautomère peut s’apparier de manière erronée à une autre base et provoquer une mutation lors de la réplication de l’ADN.
- Par exemple, la forme tautomère imino de la cytosine peut s’apparier à l’adénine au lieu de la guanine ce qui conduit à des mutations G≡C ➞ A=T (
 désamination de la cytosine).
désamination de la cytosine).
b. Les formes tautomères peuvent modifier la géométrie des paires de bases et les liaisons hydrogène, perturbant localement l’organisation de la double hélice,, en particulier si ces anomalies ne sont pas corrigées par les systèmes de réparation de l'ADN.
c. Dans certains ribozymes, notamment ceux du du groupe I, un tautomère de la guanine pourrait participer à la stabilisation de l’état de transition de la réaction catalytique. Fonctions dans les ARN structuraux et régulateurs
d. Dans les ARN structuraux ou régulateurs, les tautomères peuvent
- modifier l’appariement de bases et permettre des interactions non canoniques,
- participer à la reconnaissance moléculaire, comme dans certains riboswitches, structures d’ARN capables de lier directement des métabolites et de réguler l’expression génique.
- créer des microdomaines de stabilité ou de flexibilité dans les boucles, jonctions ou pseudonœuds des ARN.
Propriétés chimiques
1. À pH physiologique, les bases azotées sont globalement non chargées, ce qui favorise leur empilement par interactions π-π dans l’ADN.
a. Cependant, elles présentent des pKa caractéristiques, i.e. propension de certains atomes, notamment les atomes d’azote dans les bases azotées, à accepter ou à libérer un proton.
- Lors de fortes variations du pH, des changements de protonation peuvent perturber la complémentarité des bases et provoquer une dénaturation de l’ADN ou de l’ARN.
- Ces variations de pKa expliquent également certains mécanismes de catalyse acide-base observés dans les ribozymes.
b. Les mentions " protoné " ou " déprotoné " indiquent uniquement l’état d’un site azoté particulier susceptible de gagner ou perdre un proton, sans que la molécule porte nécessairement une charge nette.
| Base | Groupement concerné |
pKa | État à pH physiologique (≈7,4) |
|---|---|---|---|
| Adénine (A) | N1 (protonation possible) |
~3,5 | Déprotoné (neutre) |
| N9 (déprotonation possible) |
~9,8 | Majoritairement protoné (neutre) | |
| Guanine (G) | N1 (déprotonation) | ~9,2 | Majoritairement protoné (neutre) |
| N7 (protonation) | ~2,4 | Déprotoné | |
| Cytosine (C) | N3 (protonation) | ~4,5 | Déprotoné (neutre) |
| Uracile (U) | N3 (protonation) | ~9,5 | Déprotoné (neutre) |
| Thymine (T) | N3 (protonation) | ~10,0 | Déprotoné (neutre) |
2. Les bases, bien que polaires, présentent une tendance hydrophobe sur leur cycle aromatique, i.e. leur face plane ( effet hydrophobe sur l'agencement des baes)
Cette propriété favorise l’empilement hydrophobe à l’intérieur de la double hélice, avec les sucres-phosphates polaires exposés à l’eau.
3. Chaque base présente des donneurs et accepteurs de liaisons hydrogène spécifiques, i.e. groupes –NH, –C=O, qui conditionne la complémentarité stricte
- 2 liaisons hydrogène, A=T
- 3 liaisons hydrogène, G≡C.
Propriétés géométriques
1. La structure quasi-plane des bases azotées des acides nucléiques résulte de la délocalisation des électrons π dans les cycles, ce qui confère une rigidité conformationnelle à ces molécules ( propriétés des cycles aromatiques).
- Les bases pyrimidiques sont parfaitement planes, en raison de leur structure à un seul cycle.
- Les bases puriques, plus volumineuses par leurs deux cycles fusionnés, présentent une très légère déviation par rapport au plan, i.e. moins de 0,05 nm = 0,5 Å, ce qui reste négligeable pour l'empilement.
Les bases puriques sont considérées comme planaires à l’échelle moléculaire.
Cette planéité permet un empilement stable des bases azotées dans la double hélice
2. La longueur des cycles correspond :
- pour les pyrimidines à ≈ 0,5–0,6 nm,
- pour les purines à ≈ 0,6–0,7 nm.
a. Or, la largeur d’une paire de bases C-G ou A-T est d'environ 1,1 nm et la distance inter-base, i.e. empilement vertical dans la double hélice, est d'environ 0,34 nm.
Le diamètre de la double hélice, incluant les bases et le squelette sucre-phosphate, est d'environ 2,0–2,2 nm.
Ces dimensions sont compatibles entre paires complémentaires, ce qui assure une géométrie régulière de l’ADN en double hélice, sans distorsion majeure entre les paires A=T et G≡C.
b. Les dimensions précises des bases garantissent que seules les paires complémentaires peuvent former des liaisons hydrogène optimales, sans tension stérique, condition indispensable à la fidélité de la réplication et à la stabilité de la structure de l’ADN.
Conséquences sur l'agencement des base dans l'ADN
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesBases azotéesPentosesDifférents nucléosidesDifférents nucléotidesBiosynthèse des nucléotidesVoies des sauvetageCatabolisme des nucléotidesADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation