


Acides nucléiques
Chromatine
Code des histones : acétylation et méthylation
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques

L’acétylation et la méthylation des histones sont les modifications post-traductionnelles les plus importantes du code des histones et jouent un rôle central dans la régulation de la chromatine et de l’expression des gènes.
Parmi les nombreuses modifications post-traductionnelles des histones, l’acétylation et la méthylation sont les plus abondantes et les mieux caractérisées.
- Ces modifications affectent principalement les résidus lysine et arginine des histones de cœur, en particulier de H3 et H4, et participent à la régulation de l’organisation de la chromatine et de l’activité transcriptionnelle du génome.
- Leur étude a permis de mettre en évidence une grande partie des mécanismes moléculaires à l’origine du code des histones.
Acétylation des histones
Parmi les différentes modifications post-traductionnelles affectant les histones, l’acétylation est l’une des premières décrites et des mieux caractérisées, jouant un rôle majeur dans la régulation de la structure de la chromatine et de l’expression des gènes.
Enzymes d’acétylation et de désacétylation des histones
1. L’acétylation correspond à l'ajout d'un groupe acétyle ($\ce{-C(=O)-CH3}$) et s’effectue principalement sur les résidus lysines (K) libres des queues N-terminales des histones H3 et H4 (Post‐translational modifications of histones: Mechanisms, biological functions, and therapeutic targets 2023)
a. Cette réaction est catalysée par les histone acétyltransférases (HAT), i.e. EC 2.3.1.48 :
$\ce{L-lysyl-[histone] + acétyl-CoA}$ $\longrightarrow$ $\ce{N(6)-acétyl-L-lysyl-[histone] + CoA + H+}$
b. Les HAT, qui sont des writers dans le cadre du code des histones, sont classées en plusieurs familles.
- Les GNAT (GCN5-related N-acetyltransferases) incluent GCN5 et PCAF et sont souvent associées à des complexes activateurs de la transcription.
- La famille MYST, comprenant MOZ, Ybf2/Sas3, Sas2 et Tip60, est impliquée dans la régulation de la transcription, la réparation de l’ADN et la régulation de la croissance cellulaire, en interphase et particulièrement en phase G1.
- CBP/p300, coactivateurs transcriptionnels majeurs, possèdent une activité HAT à large spectre, ciblant de nombreux résidus d’histones et de protéines non histones.
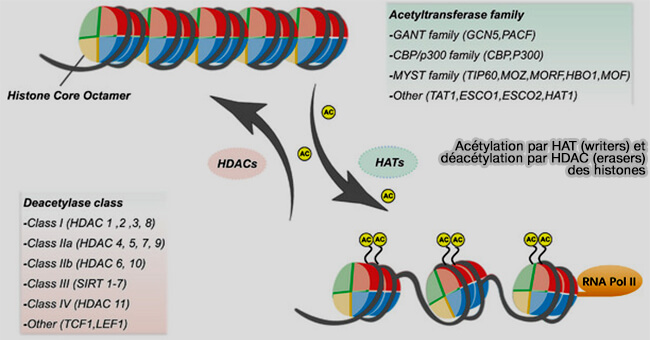
(Figure : vetopsy.fr d'après Liu et coll)
2. L'acétylation est réversible grâce aux histone déacétylases (HDAC), i.e. EC 3.5.1.98, qui retirent le groupement acétyl :
$\ce{N(6)-acétyl-L-lysyl-[histone] + H2O}$ $\longrightarrow$ $\ce{L-lysyl-[histone] + acétate}$
Les HDAC, qui sont des erasers dans le cadre du code des histones, sont regroupées en plusieurs classes, souvent en fonction de leur homologie avec les enzymes de levure.
- La classe I comprend HDAC1, 2, 3, 8, principalement nucléaires, impliquées dans la répression transcriptionnelle.
- La classe II comprend HDAC4, 5, 6, 7, 9, 10, capables de faire la navette entre le noyau et le cytoplasme, a un rôle dans la différenciation cellulaire et la signalisation.
- La classe III (sirtuines), composée de SIRT1 à SIRT7, enzymes NAD+-dépendantes, est impliquée dans la régulation épigénétique, le métabolisme et le vieillissement.
- La classe IV, comprenant le seul HDAC11, possède des caractéristiques intermédiaires entre les classes I et II.

L’équilibre entre l’activité des HAT et des HDAC détermine l’état d’acétylation des histones et influence directement la compaction de la chromatine et la régulation transcriptionnelle, les HAT favorisant l’expression des gènes, tandis que les HDAC répriment certains gènes pour permettre une progression ordonnée.
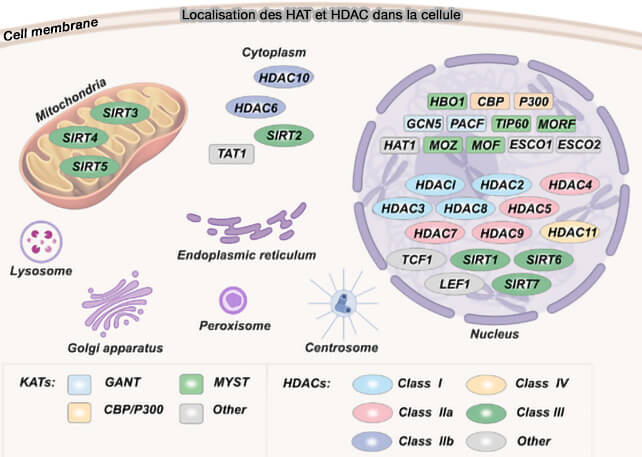
(Figure : vetopsy.fr d'après Liu et coll)
Effets structuraux de l’acétylation sur la chromatine
L’acétylation des lysines modifie directement les propriétés physico-chimiques des histones.
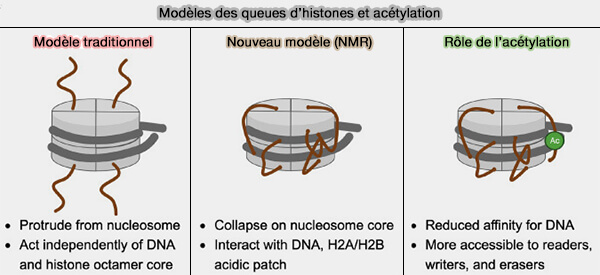
(Figure : vetopsy.fr d'après Weinzapfel et coll)
1. L’ajout d’un groupe acétyle neutralise la charge positive du groupement ε-amine de la lysine (Beyond the tail: the consequence of context in histone post-translational modification and chromatin research 2024).
- Cette neutralisation réduit l’attraction électrostatique entre les queues d'histones et le squelette phosphate de l’ADN, chargé négativement.
- La diminution de ces interactions affaiblit également les contacts entre nucléosomes voisins, notamment ceux impliquant les queues N-terminales des histones H3 et H4.
2. Ces modifications entraînent un relâchement de l’organisation chromatinienne, généralement associé à l’euchromatine et à une augmentation de l’accessibilité de l’ADN.
Reconnaissance des lysines acétylées et fonctions cellulaires
1. Les lysines acétylées constituent des sites de reconnaissance spécifiques pour des protéines lectrices, readers dans le cadre du code des histones, capables d’interpréter ces modifications.
- De nombreuses protéines, readers dans le cadre du code des histones, possèdent des bromodomaines, domaines structuraux spécialisés qui reconnaissent et se lient aux lysines acétylées présentes sur les queues d'histones.
- La fixation de ces protéines favorise le recrutement de divers complexes chromatiniens impliqués dans la régulation de la transcription, notamment des complexes de remodelage de la chromatine et des coactivateurs transcriptionnels.
L'acétylation favorise la transcription active en permettant la liaison des facteurs de transcription et de la machinerie transcriptionnelle.
2. L’accumulation de plusieurs lysines acétylées sur H3 et H4 est généralement corrélée à des régions de transcription intense et certaines marques sont particulièrement caractéristiques.
- H3K9ac et H3K14ac sont des marques classiques d’activation transcriptionnelle locale, souvent présentes au niveau des promoteurs.
- H4K16ac joue un rôle clé dans le relâchement de la fibre de chromatine en inhibant sa compaction et joue un rôle majeur dans l’organisation globale du génome et dans la régulation de l’expression de larges domaines génomiques, régions étendues d’un chromosome ayant un état chromatinien homogène (
 organisation tridimensionnelle de la chromatine).
organisation tridimensionnelle de la chromatine).
3. L’acétylation des histones facilite également d’autres processus nucléaires en rendant la chromatine plus accessible aux complexes enzymatiques impliqués dans :
- l’initiation de la réplication de l'ADN,
- la réparation de l’ADN,
4. Des altérations du contrôle de l’acétylation des histones sont associées à diverses pathologies, notamment certains cancers et des et des processus liés au vieillissement cellulaire.
a. Des mutations ou des réarrangements affectant des histone acétyltransférases comme CBP/p300 ou MOZ ont été identifiés dans plusieurs leucémies et tumeurs solides.
- À l’inverse, une activité excessive des histone désacétylases peut maintenir des gènes suppresseurs de tumeurs dans un état réprimé, contribuant à la progression tumorale.
- Ces observations ont conduit au développement d’inhibiteurs d’HDAC utilisés comme agents thérapeutiques dans certains cancers, par exemple le vorinostat (SAHA) et la romidepsine, utilisés dans le traitement de certains lymphomes T cutanés.
b. Les sirtuines (classe III des HDAC) jouent un rôle important dans le contrôle du métabolisme énergétique, de la réponse au stress oxydatif et de certains mécanismes associés au vieillissement.
3. Les acétylations majeures sont résumées dans le tableau suivant.
| Histone | Résidu acétylé | Rôle principal | Exemples biologiques |
|---|---|---|---|
| H3 | K9 | Activation transcriptionnelle |
Gènes de différenciation |
| K14 | Activation transcriptionnelle |
|
|
| K18, K23 | Activation transcriptionnelle |
Régulation transcriptionnelle |
|
| K27 | Activation transcriptionnelle |
|
|
| K56 |
|
|
|
| H4 | K5, K8, K12, K16 | Ouverture chromatine |
Transcription active |
| H2A | K5, K9, K13 | Activation transcriptionnelle |
Réponse aux signaux cellulaires |
| H2B | K5, K12, K15, K20 | Activation transcriptionnelle |
Organisation de la chromatine |
Méthylation des histones
La méthylation des histones correspond à l’ajout de groupes méthyle sur certains résidus des histones et constitue l’une des modifications majeures du code des histones impliquées dans la régulation de l’activité chromatinienne.
Enzymes de méthylation et de déméthylation des histones
1. La méthylation des histones s’effectue sur des lysines (K) ou des arginines (R) des histones et peut consister en l'ajout d'un à trois groupes méthyle ($\ce{-CH3}$), et appelées me1, me2 ou me3.
a. Cette réaction est catalysée par les histone méthyltransférases (HMT), i.e. EC 2.1.1.-, writers qui utilisent généralement la S-adénosylméthionine (SAM) comme donneur de groupe méthyle.
$\ce{[histone]-Lys + S-adénosyl-L-méthionine}$ $\longrightarrow$
$\ce{[histone]-Lys-CH3 + S-adénosyl-L-homocystéine}$
Remarque : en pratique, beaucoup d’enzymes d’histones ont aujourd’hui des EC spécifiques par résidu et par état de méthylation dans la classe EC 2.1.1-.
- Pour les histone-lysine méthyltransférases, l’EC général historique est EC 2.1.1.43, maintenant .354,
- Pour les protein-arginine methyltransferases (PRMT), l’EC général est EC 2.1.1.23., maintenant .319, .320, .321, .322.
- En pratique, beaucoup d’enzymes d’histones ont aujourd’hui des EC spécifiques par résidu et par état de méthylation dans la classe EC 2.1.1.-, par exemple pour H3K4 (.354), H3K9 (.355) ou H3K27 (.356).
b. Les lysines peuvent être mono-, di- ou triméthylées, tandis que les arginines peuvent être mono- ou diméthylées, ces dernières pouvant être symétriques ou asymétriques.
- Pour les histone-lysine méthyltransférases, l’EC général historique est EC 2.1.1.43.
- Pour les protein-arginine methyltransferases (PRMT), l’EC général est EC 2.1.1.23.
- En pratique, beaucoup d’enzymes d’histones ont aujourd’hui des EC spécifiques par résidu et par état de méthylation dans la classe EC 2.1.1.-, par exemple pour H3K4, H3K9 ou H3K27.
2. Cette réaction est réversible grâce aux histone déméthylases (HDM), EC 1.14.11.27, erasers capables d’éliminer les groupes méthyle présents sur les lysines ou les arginines des histones :
Les déméthylases appartiennent principalement à deux grandes familles :
- les enzymes LSD (lysine specific demethylase) dépendantes du FAD, EC 1.5.1.16, qui agissent sur me2 et me1,
$\ce{N(6)-methyl-L-lysyl(36)-[histone H3] + O2}$ $\leftrightharpoons$
$\ce{L-lysyl(36)-[histone H3] + formaldehyde + H2O2}$
- les déméthylases JmjC, dépendantes du Fe++ et du 2-oxoglutarate, EC 1.14.11.-, qui agissent sur me3 (.66 à .69), me2 (.27 et .65) et me1, par exemple pour me1,
$\ce{N(6)-methyl-L-lysyl(36)-[histone H3] + 2-oxoglutarate + O2}$ $\leftrightharpoons$
$\ce{L-lysyl(36)-[histone H3] + formaldehyde + succinate + CO2}$
Effets structuraux de la méthylation sur la chromatine
1. La méthylation des histones modifie les propriétés fonctionnelles de la chromatine mais, contrairement à l’acétylation, elle n’altère pas directement la charge électrique des résidus modifiés.
La méthylation n’entraîne donc pas directement un relâchement ou une compaction de la chromatine par modification des charges.
2. Les effets structuraux de la méthylation dépendent principalement du résidu modifié et du degré de méthylation.
Les différentes formes de méthylation constituent surtout des signaux moléculaires permettant le recrutement de protéines lectrices, qui déterminent ensuite l’état fonctionnel des régions chromatiniennes.
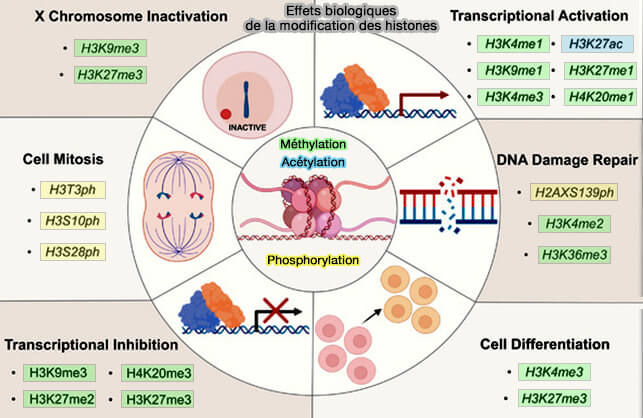
(Figure : vetopsy.fr d'après Liu et coll)
Reconnaissance des lysines et des arginines méthylées
Les résidus lysines (K) et arginines (R) méthylés des histones constituent des sites de reconnaissance pour des protéines lectrices, capables d’interpréter ces marques et de recruter divers complexes chromatiniens.
1. Ces protéines possèdent des domaines structuraux spécialisés capables de reconnaître certaines méthylations spécifiques des histones.
Les domaines de lecture des histones méthylées (readers) sont étudiés dans un chapitre spécifique.
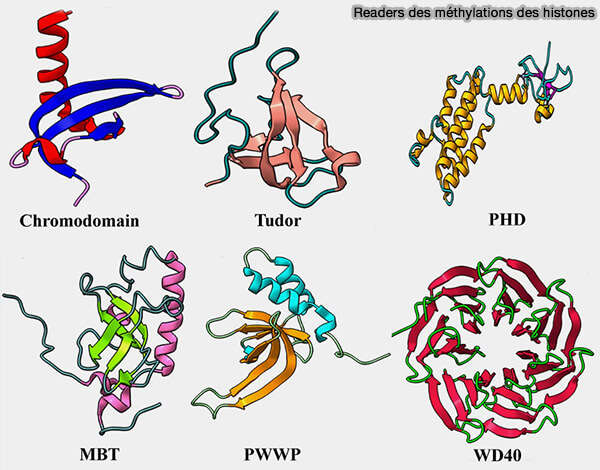
(Figure : vetopsy.fr d'après Sehrawat et coll)
2. La reconnaissance de ces marques permet le recrutement de complexes de régulation de la chromatine.
Remarque : certaines modifications d’histones peuvent être antagonistes, comme l’acétylation d’une lysine qui empêche généralement sa méthylation.
Cet antagonisme contribue à l’établissement d’un équilibre dynamique entre domaines euchromatiniens, souvent acétylés et transcriptionnellement actifs, et domaines hétérochromatiniens, plus fréquemment associés à certaines méthylations répressives comme H3K9me3.
Méthylations majeures des histones
Les modifications par méthylation concernent principalement les histones de cœur H3 et H4, qui portent la majorité des sites méthylés étudiés.
- Les histones H2A/H2B présentent également quelques résidus méthylés, mais ces modifications sont beaucoup moins caractérisées.
- L’histone H1, histone de liaison, peut aussi porter certaines marques de méthylation, mais elles restent rares et encore peu étudiées.
1. Ces modifications, selon le résidu modifié et le degré de méthylation, peuvent être associées :
- à la régulation de la transcription, soit par l’activation transcriptionnelle, soit par la répression stable de l’expression génique,
- à la différenciation cellulaire,
- à l’organisation des domaines d'hétérochromatine,
- au maintien d’états épigénétiques au cours des divisions cellulaires, conservation ou rétablissement après la réplication de l’ADN et transmissions aux cellules filles, permettant ainsi de maintenir le même état d’expression des gènes au cours des divisions cellulaires, sans modifier la séquence de l’ADN ( épigénétique et régulation épigénétique).
2. Parmi les nombreuses méthylations identifiées, certaines marques sont particulièrement bien caractérisées et jouent un rôle central dans l’organisation fonctionnelle de la chromatine.
- H3K4me3 est une marque enrichie au niveau des promoteurs des gènes actifs et associée à l’initiation de la transcription.
- H3K9me3 est une marque répressive caractéristique de l’hétérochromatine constitutive, impliquée dans le silence transcriptionnel stable et le recrutement de HP1 (Heterochromatin Protein 1).
- H3K27me3 est déposée par le complexe PRC2 et participe à la répression réversible de nombreux gènes du développement dans l’hétérochromatine facultative.
- H3K36me3 est associée au corps des gènes activement transcrits, où elle accompagne l’élongation transcriptionnelle, la fidélité de l’épissage et certains mécanismes de réparation de l’ADN.
- H3K79me2/3 est localisée dans le globule de l’histone H3 et intervient dans l’élongation transcriptionnelle ainsi que dans la réponse aux dommages de l’ADN (DDR).
- H4K20me1/2/3 participe selon son degré de méthylation à la réplication de l’ADN, à la compaction de la chromatine, à la réparation de l’ADN et à l’organisation de l’hétérochromatine.
3. Ces méthylations majeures sont résumées dans le tableau suivant.
| Histone | Résidu méthylé | Type de méthylation | Rôle fonctionnel | Exemples biologiques |
|---|---|---|---|---|
| Marques associées à la chromatine active | ||||
| H3 | K4 | me1, me2, me3 | Activation transcriptionnelle |
|
| K36 | me1, me2, me3 | Élongation transcriptionnelle | Gènes fortement exprimés | |
| K79 | me2, me3 |
|
|
|
| Marques associées à la chromatine réprimée | ||||
| H3 | K9 | me1, me2, me3 |
|
Silencing séquences répétées |
| K27 | me1, me2, me3 | Répression transcriptionnelle (principalement H3K27me3) via PRC2 |
|
|
| H4 | K20 | me1, me2, me3 | Organisation et compaction de la chromatine |
Hétérochromatine |
| Marques de régulation fine | ||||
| H3 | R2, R8, R17, R26 | me1, me2 (sym/asym) | Régulation transcriptionnelle fine |
Réponse aux signaux cellulaires |
| H1 | K26 (H1.4) | me1, me2 | Répression transcriptionnelle avec H3K9me2/3 |
Doamines hétérochromatiniens |
Remarque : malgré sa complexité, le tableau est simplifié car les différents degrés de méthylation (me1, me2, me3) peuvent avoir des fonctions distinctes selon le résidu concerné.
- H3K4me3 intervient dans les promoteurs actifs, H3K4me2 dans les régions actives intermédiaires et H3K4me1 comme enhancer.
- H3K36me3 intervient dans l'élongation transcriptionnelle, H3K36me2 dans l'organisation chromatinienne et H3K36me1 a un rôle plus variable.
Propagation de l’hétérochromatine : SUV39 et HP1
Certaines méthylations d’histones participent à la propagation locale de l’hétérochromatine.
1. L’établissement de ces domaines repose notamment sur la méthylation de la lysine 9 de l’histone H3 (H3K9me2/3), catalysée par des histone méthyltransférases de la famille SUV39H1/2 (Structure, Activity and Function of the Suv39h1 and Suv39h2 Protein Lysine Methyltransferases 2021).
- Cette réaction est catalysée par les histone-lysine méthyltransférases SUV39H1 et SUV39H2, qui possèdent un domaine SET responsable du transfert de groupes méthyle depuis la S-adénosyl-méthionine (SAM) vers la lysine 9 de H3.
Ces deux enzymes sont très proches, mais montrent des profils d’expression et des rôles partiellement distincts selon les tissus et les stades du développement.
- La lysine peut être mono-, di- ou triméthylée, mais H3K9me3 est la marque typique de l’hétérochromatine constitutive.
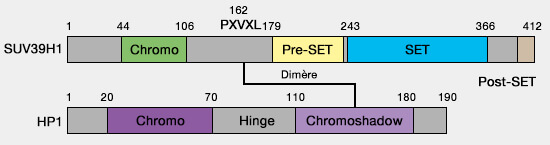
(Figure : vetopsy.fr)
2. La marque H3K9me2/3 est reconnue par HP1 (Heterochromatin Protein 1) qui possède deux domaines structuraux (Division of labor between the chromodomains of HP1 and Suv39 methylase enables coordination of heterochromatin spread 2014) :
- un chromodomaine (CD) N-terminal qui reconnaît spécifiquement H3K9me2/3 sur la queue N-terminale de l’histone H3 et l’insère dans une poche aromatique, ce qui stabilise la liaison au nucléosome,
- un domaine chromoshadow (CSD) C-terminal responsable de la dimérisation de HP1 et du recrutement de protéines partenaires.
a. HP1 existe chez les vertébrés sous trois isoformes qui possèdent la même organisation structurale et reconnaissent la marque H3K9me2/3, mais diffèrent par leur localisation chromatinienne et leurs fonctions dans l’organisation de la chromatine et la régulation transcriptionnelle (HP1: Heterochromatin binding proteins working the genome 2011 et Mammalian HP1 Isoforms Have Specific Roles in Heterochromatin Structure and Organization 2017) :
- HP1α/CBX5, principalement associée à l’hétérochromatine constitutive dans les régions péricentromériques,
- HP1β/CBX1, mpliquée notamment dans la stabilité génomique et la réparation de l’ADN,
- HP1γ/CBX3, capable d’intervenir dans des contextes chromatiniens plus dynamiques, y compris dans certaines régions euchromatiniennes associées à la régulation transcriptionnelle.
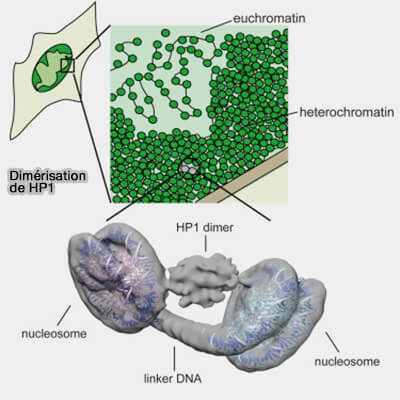
(Figure : vetopsy.fr d'après Mashida et coll)
b. Outre la phosphorylation bien établie de l'histone H3 Ser10 qui module la liaison de HP1 à la chromatine, plusieurs études ont récemment souligné l'importance des modifications post-traductionnelles de HP1 et des caractéristiques épigénétiques supplémentaires pour la modulation de la capacité de liaison de HP1 à la chromatine et la formation d'hétérochromatine (How HP1 Post-Translational Modifications Regulate Heterochromatin Formation and Maintenance 2020).
3. Le chromoshadow domain permet à deux molécules de HP1 de former un dimère stable, ce qui a plusieurs conséquences structurales importantes (Structural Basis of Heterochromatin Formation by Human HP1 2018) :
- chaque monomère peut se fixer à un nucléosome portant H3K9me3,
- le dimère peut relier deux nucléosomes voisins, ce qui favorise l’agrégation locale de nucléosomes méthylés.
4. Une fois le dimère formé, l’interface entre les deux domaines chromoshadow crée une poche de liaison capable de reconnaître des protéines contenant un motif peptidique spécifique (souvent PXVXL), présent dans différentes protéines associées à l’hétérochromatine, dont SUV39H.
- Ainsi, HP1 peut recruter SUV39H sur la chromatine déjà méthylée.
- Une fois recrutée, SUV39H méthyle les nucléosomes voisins, déposant de nouvelles marques H3K9me2/3.
- Ces nouvelles marques deviennent à leur tour des sites de fixation pour HP1, ce qui permet la fixation de nouveaux dimères de HP1, le recrutement supplémentaire de SUV39H et la méthylation de nucléosomes adjacents.
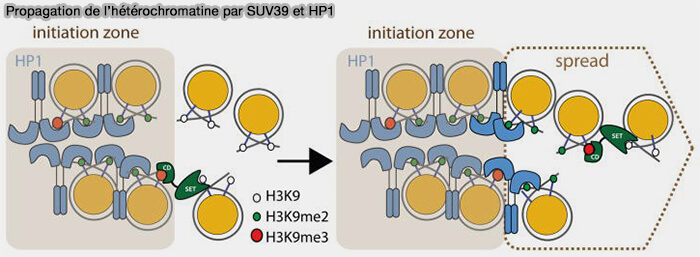
(Figure : vetopsy.fr d'après Al-Sady et coll)
Ce mécanisme crée une boucle d’auto-renforcement, souvent appelée mécanisme " read-write ", SUV39H " écrit " la marque H3K9me, HP1 " lit " cette marque et recrute SUV39H, permettant l’extension progressive des domaines riches en H3K9me3 le long de la chromatine.
Il en résulte la formation d’un domaine d’hétérochromatine transcriptionnellement silencieux, typiquement observé dans les centromères, les régions péricentromériques, certaines régions répétées du génome.
5. En outre, sans ces régions hétérochromatiniennes établies, l’histone de liaison H1 peut contribuer à la stabilisation de la structure compacte.
Certaines modifications de H1, notamment H1.4K26me1/2, sont associées à ces domaines répressifs et participent au renforcement de l’organisation de l’hétérochromatine.
Remarque : l’acétylation du résidu H3K9 peut empêcher sa méthylation, freinant l’expansion du domaine hétérochromatinien et contribuant à l’établissement d’un équilibre dynamique entre euchromatine et hétérochromatine dans le génome.
Autres modifications post-traductionnelles des histones
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNChromatineStructure dynamique de la chromatineNucléosomesHistonesMembres des histonesCode des histonesChromosomesStructure des chromosomesCentromèreTélomèreProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation