


Acides nucléiques
ADN : épigénétique
Mécanismes
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

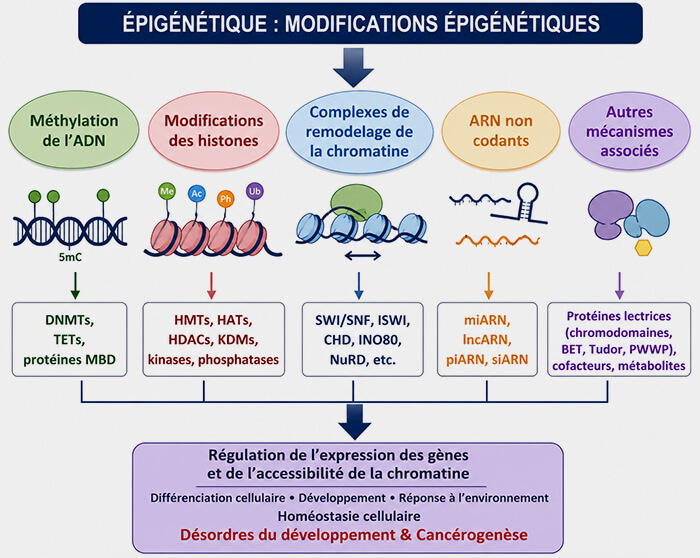
L'épigénétique désigne l'ensemble des mécanismes moléculaires qui modulent durablement l'expression des gènes sans modifier la séquence nucléotidique de l'ADN.
Les principaux mécanismes épigénétiques reposent sur :
- des modifications chimiques de l'ADN et des histones,
- le remodelage de la chromatine,
- l'action de certains ARN non codants.
(Figure : vetopsy.fr)
En modifiant l'accessibilité de l'ADN aux facteurs de transcription et à la machinerie transcriptionnelle, ces mécanismes déterminent quels gènes sont exprimés, réprimés ou susceptibles d'être activés selon le type cellulaire, le stade du développement ou les conditions environnementales.
L'épigénétique joue un rôle essentiel dans :
- la différenciation cellulaire,
- le développement embryonnaire,
- le maintien de l'identité des cellules,
- l'adaptation aux variations de l'environnement,
- de nombreux processus physiologiques et pathologiques.
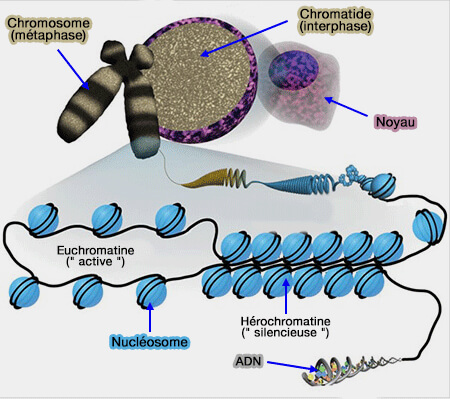
Organisation de la chromatine
(Figure : vetopsy.fr d'après Sha-Boyer)
{kind=link}
Les mécanismes épigénétiques s'exercent principalement au niveau de la chromatine, complexe constitué d'ADN associé aux histones et à de nombreuses protéines non histones.
1. Selon son degré de compaction, la chromatine adopte des états plus ou moins accessibles aux facteurs de transcription et aux différentes protéines impliquées dans l'expression génique.
- L'euchromatine peu condensée favorise généralement la transcription.
- L'hétérochromatine fortement condensée est le plus souvent associée à une répression transcriptionnelle.
2. Les modifications épigénétiques agissent en modifiant localement cette organisation de la chromatine, rendant certaines régions du génome plus accessibles ou, au contraire, moins accessibles à la machinerie transcriptionnelle.
Principaux mécanismes épigénétiques
Les principaux mécanismes comprennent :
- les marques épigénétiques, notamment méthylation de l'ADN et modifications post-traductionnelles et variants des histones,
- le remodelage de la chromatine,
- l'action de certains ARN non codants.

Ces différents mécanismes n'agissent pas de manière indépendante, mais interagissent étroitement pour établir, maintenir ou modifier l'état transcriptionnel des gènes en fonction du type cellulaire, du stade du développement ou des conditions environnementales.
Marques épigénétiques
Les marques épigénétiques sont des modifications chimiques réversibles de l'ADN ou des histones qui modulent l'expression des gènes sans modifier la séquence nucléotidique de l'ADN.
- Elles constituent le principal support moléculaire de l'information épigénétique.
- Associées à l'organisation de la chromatine et à l'action de certains ARN non codants (ncRNA), elles contribuent à établir, à maintenir et à modifier durablement les états d'expression génique.
Méthylation de l’ADN
Chez les eucaryotes, la méthylation de l'ADN est l'un des principaux mécanismes épigénétiques de régulation de l'expression génique.
Selon sa localisation dans le génome, elle est généralement associée à une répression transcriptionnelle, tout en participant à d'autres fonctions biologiques. (A comprehensive review of structural insights into DNA methylation maintenance 2025).
Principe général
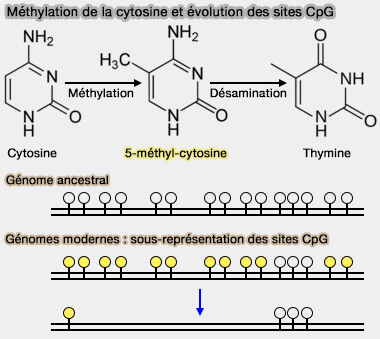
1. La méthylation de l'aDN correspond à l'ajout d'un groupe méthyle (-CH3) sur certaines bases de l'ADN, sans modifier sa séquence nucléotidique.
(Figure : vetopsy.fr)
a. Chez les vertébrés, cette modification concerne principalement la cytosine (C) des dinucléotides CpG (cytosine-phosphate-guanine), convertie en 5-méthylcytosine (5mC), l'une des principales marques épigénétiques de l'ADN.
- La 5-hydroxyméthylcytosine (5hmC), produite par oxydation de la 5mC par les enzymes TET, est particulièrement abondante dans certains tissus, notamment le cerveau, où elle possède également des fonctions régulatrices propres.
- Les dérivés 5-formylcytosine (5fC) et 5-carboxylcytosine (5caC) sont beaucoup plus rares et interviennent principalement comme intermédiaires de la déméthylation active de l'ADN.
Remarque : contrairement à la 5-méthylcytosine, la N6-méthyladénine (6mA) est une modification majeure de l'ADN chez les bactéries, tandis que sa présence et son rôle épigénétique chez les mammifères demeurent débattus (N6-methyladenine is incorporated into mammalian genome by DNA polymerase 2020).
b. Les dinucléotides CpG sont répartis dans l'ensemble du génome, mais se concentrent dans certaines régions appelées îlots CpG, fréquemment localisées au niveau des promoteurs de nombreux gènes ( cf plus bas).
cf plus bas).
2. La méthylation est catalysée par les ADN méthyltransférases (DNMT).
a. Les enzymes DNMT3A et DNMT3B établissent de nouvelles marques de méthylation (de novo) en transférant un groupe méthyle fourni par la S-adénosylméthionine (SAM) sur les cytosines des sites CpG.,
b. DNMT1 assure principalement le maintien de ces marques lors de la réplication de l'ADN (Two distinct modes of DNMT1 recruitment ensure stable maintenance DNA methylation 2020).
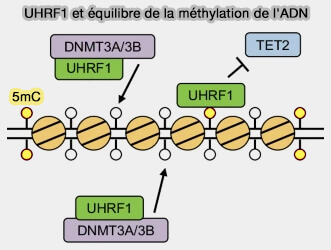
- Au début de la réplication, la protéine UHRF1 (Ubiquitin-like with PHD and RING Finger Domains 1) reconnaît les sites CpG hémi-méthylés et recrute DNMT1 grâce à l'ubiquitinylation de PAF15 (PCNA-associated factor 15), une protéine associée au complexe de réplication.
- Après le réassemblage des nucléosomes, UHRF1 recrute principalement DNMT1 par l'ubiquitinylation de l'histone H3.
- Dans les deux cas, DNMT1 méthyle alors les cytosines correspondantes du brin néosynthétisé, rétablissant la méthylation des sites CpG et assurant ainsi la transmission fidèle des profils de méthylation aux cellules filles.
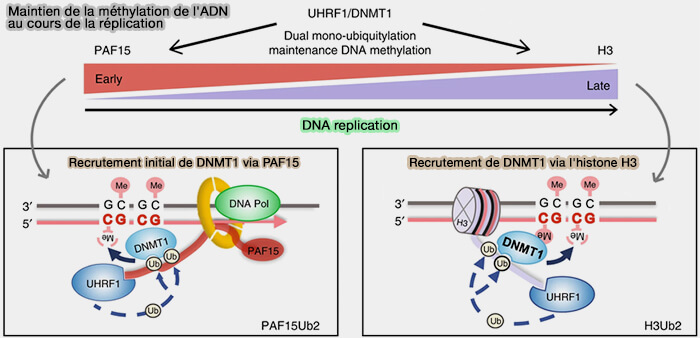
(Figure : vetopsy.fr modifiée d'après Vuu et coll)
3. À l'inverse, la déméthylation peut être :
- passive, par absence de maintien des marques au cours des divisions cellulaires,
- active grâce aux enzymes TET1/2/3, qui oxydent successivement la 5-méthylcytosine en 5-hydroxyméthylcytosine (5hmC), puis en 5-formylcytosine (5fC) et en 5-carboxylcytosine (5caC) qui sont ensuite remplacées par une cytosine non méthylée grâce aux mécanismes de réparation par excision de base (BER).
(Figure : vetopsy.fr d'après Yamagushi et coll)
Remarque : outre son rôle dans le maintien de la méthylation de l'ADN par le recrutement de DNMT1, UHRF1 participe également à l'homéostasie de la méthylation (Non-canonical functions of UHRF1 maintain DNA methylation homeostasis in cancer cells 2024) :
- en favorisant, selon le contexte cellulaire, l'activité des méthyltransférases DNMT3A et DNMT3B,
- en limitant celle des enzymes TET, contribuant ainsi à l'équilibre entre méthylation de novo, maintien et déméthylation.
Îlots CpG
Les îlots CpG correspondent à des régions de l'ADN particulièrement riches en sites CpG, généralement localisées au niveau des promoteurs de nombreux gènes chez les vertébrés (Intragenic CpG Islands and Their Impact on Gene Regulation 2022).
1. Un site CpG correspond à un dinucléotide dans lequel une cytosine (C) est suivie directement d’une guanine (G) sur le même brin d'ADN, les deux nucléotides étant reliés par un groupement phosphate (d’où le " p " de CpG, pour cytosine-phosphate-guanine).
- Cette notation permet de distinguer les dinucléotides CpG des paires de bases C-G reliant les deux brins de la double hélice.
- Chez les eucaryotes, les cytosines des sites CpG peuvent être méthylées en position 5 du cycle pyrimidique pour former la 5-méthylcytosine (5mC), l'une des principales marques épigénétiques de l'ADN.
(Figure : vetopsy.fr modifiée d'après Cain et coll)
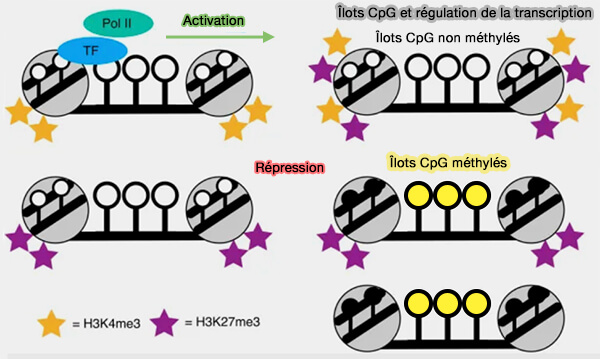
2. Alors que la majorité des sites CpG situés en dehors des îlots CpG sont méthylés, les îlots CpG sont associés aux promoteurs des gènes exprimés et restent généralement non méthylés ou faiblement méthylés.
- Cet état favorise le maintien d'une chromatine accessible, permettant la fixation des facteurs de transcription et le recrutement de l'ARN polymérase II.
- Les promoteurs contenant des îlots CpG sont particulièrement fréquents dans les gènes constitutivement exprimés (housekeeping genes) ainsi que dans de nombreux gènes impliqués dans les fonctions cellulaires fondamentales.
3. À l'inverse, la méthylation des cytosines des îlots CpG localisés dans les promoteurs est généralement associée à une répression transcriptionnelle.
Conséquences de la méthylation de l'ADN
1. La méthylation de l'ADN favorise généralement un état transcriptionnel réprimé par plusieurs mécanismes complémentaires :
- empêcher la fixation de certains facteurs de transcription,
- recruter des protéines reconnaissant l'ADN méthylé,
- favoriser la condensation locale de la chromatine,
- stabiliser des états transcriptionnels silencieux.
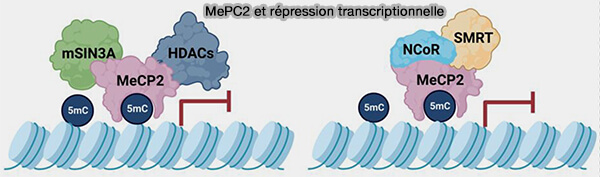
2. Les principales protéines reconnaissant l'ADN méthylé appartiennent à la famille des protéines à domaine MBD (Methyl-CpG-binding domain), notamment MeCP2, MBD1 et MBD2 qui recrutent fréquemment (Methyl-CpG-Binding Domain Proteins: Readers of the Epigenome 2015 et MeCP2 Is an Epigenetic Factor That Links DNA Methylation with Brain Metabolism 2023) :
- différents corépresseurs transcriptionnels, notamment les complexes mSIN3A, NCoR ou SMRT, qui favorisent à leur tour le recrutement (Complete Profiling of Methyl-CpG-Binding Domains for Combinations of Cytosine Modifications at CpG Dinucleotides Reveals Differential Read-out in Normal and Rett-Associated States 2020),
- des histones désacétylases (HDAC),
- des complexes de remodelage de la chromatine,
- d'autres protéines répressives, renforçant ainsi la répression de l'expression génique des histones désacétylases (HDAC).
(Figure : vetopsy.fr modifiée d'après Vuu et coll)
Remarque : chez certains types cellulaires, notamment les neurones, MeCP2 peut également être associé à des régions enrichies en 5hmC et participer à l'activation de certains gènes.
3. La méthylation de l'ADN intervient dans de nombreux processus biologiques, notamment :
- la différenciation cellulaire, en participant à l'établissement et au maintien des profils d'expression génique caractéristiques des différents types cellulaires (neurones, cellules musculaires, hépatocytes...),
- le développement embryonnaire, en contrôlant l'activation et la répression successives de nombreux gènes au cours de la formation des différents tissus.
- l'empreinte génomique, qui correspond à un mécanisme épigénétique dans lequel certains gènes sont exprimés uniquement à partir de l'allèle maternel ou de l'allèle paternel, l'autre allèle étant maintenu silencieux principalement par la méthylation de l'ADN (syndromes de Prader-Willi, d'Angelman et de Beckwith-Wiedemann…)
- l'inactivation du chromosome X, en contribuant au maintien de l'état silencieux du chromosome X inactif, après son initiation par le lncARN XIST,
- la stabilité du génome, en réprimant notamment l'expression des éléments transposables (transposons) et de nombreuses séquences répétées.
4. La méthylation de l'ADN intervient également dans l'organisation de certaines régions chromosomiques spécialisées.
Au niveau des centromères, la distribution de la méthylation des CpG contribue à distinguer la chromatine centromérique active, contenant CENP-A et le complexe CCAN, de l'hétérochromatine péricentromérique plus condensée.
Modifications post-traductionnelles et variants des histones
Avec la méthylation de l'ADN, les modifications post-traductionnelles des histones constituent les principales marques épigénétiques.
Les variants d'histones représentent un autre mécanisme épigénétique majeur contribuant à l'organisation et au fonctionnement de la chromatine (membres des histones et variants).
1. Les histones peuvent subir de nombreuses modifications post-traductionnelles, telles que :
- l'acétylation,
- la méthylation,
- la phosphorylation,
- l'ubiquitination ou la sumoylation.
a. Ces modifications modulent la compaction de la chromatine et le recrutement de protéines régulatrices, contribuant ainsi à l'activation ou à la répression de l'expression génique ( code des histones).
(Figure : vetopsy.fr d'après Malone et coll)
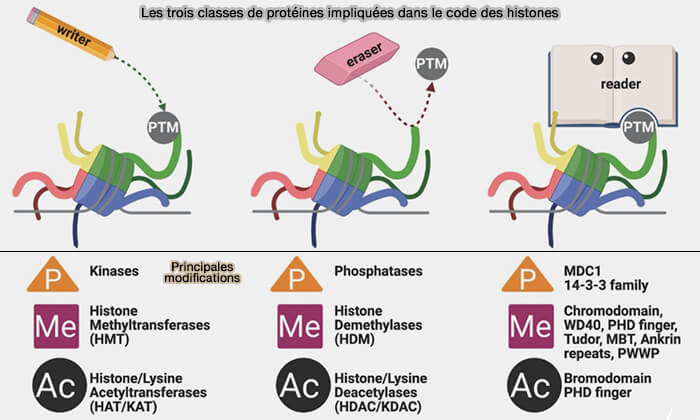
b. Ces modifications sont établies, éliminées et interprétées par trois grandes catégories de protéines intervenant dans le code des histones :
- des protéines writers, qui ajoutent les modifications,
- des protéines erasers, qui les retirent,
- des protéines readers, qui reconnaissent ces marques et recrutent les différents complexes responsables de leurs effets biologiques.
2. Les histones canoniques peuvent également être remplacées par des variants présentant des propriétés structurales et fonctionnelles spécifiques.
Parmi les principaux variants :
- H2A.Z et H3.3 participent à la régulation de l'expression génique en favorisant l'établissement d'états chromatiniens dynamiques,
- H2A.X intervient dans la réponse aux lésions de l'ADN, notamment après sa phosphorylation en γH2AX lors des cassures double brin,
- CENP-A, variant de l'histone H3, est indispensable à l'organisation et au fonctionnement des centromères,
- macroH2A est associé à des régions de chromatine transcriptionnellement réprimées, notamment au chromosome X inactif, et contribue à la mémoire épigénétique.
Autres mécanismes épigénétiques
D'autres mécanismes contribuent à la régulation épigénétique sans constituer à proprement parler des marques épigénétiques.
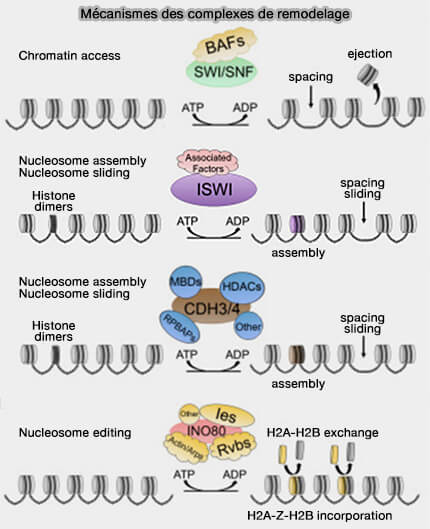
(Figure : vetopsy.fr d'après Magaña-Acosta et coll)
Ils agissent notamment en modifiant l'organisation de la chromatine ou en recrutant des complexes régulateurs sur des régions spécifiques du génome.
1. Des complexes de remodelage ATP-dépendants modifient la position, la composition ou l'organisation des nucléosomes afin de moduler l'accessibilité de l'ADN à la machinerie transcriptionnelle.
a. Ces complexes participent ainsi à la transition entre :
- l'euchromatine, chromatine ouverte généralement associée à des gènes actifs,
- l'hétérochromatine, le plus souvent associée au silence génique.
b. Les principales familles de complexes de remodelage comprennent les complexes SWI/SNF, ISWI, CHD et INO80.
Elles interviennent notamment dans le repositionnement, l'éjection, le remplacement ou la restructuration des nucléosomes en fonction du contexte cellulaire ( complexes du remodelage de la chromatine).
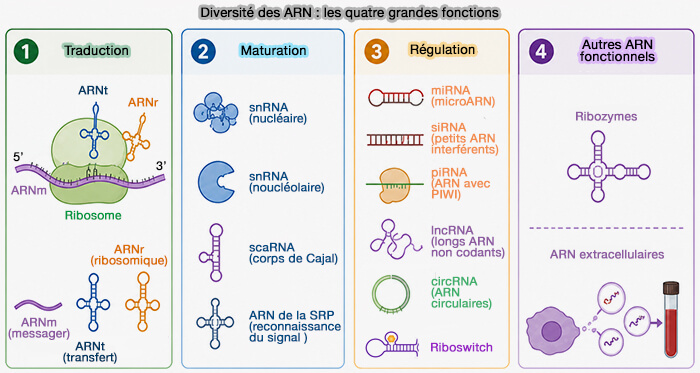
2. Plusieurs ARN non codants participent à la mise en place, au recrutement ou au maintien des mécanismes épigénétiques en interagissant avec l'ADN, les histones ou différents complexes protéiques régulateurs.
- Les ARN longs non codants (lncARN) servent souvent de molécules guides ou d'échafaudage (scaffold) en recrutant des complexes de remodelage de la chromatine ou de modification des histones sur des régions spécifiques du génome, comme XIST lors de l'inactivation du chromosome X.
- Les pRNA (promoter-associated RNA) sont des ARN non codants transcrits à partir des régions intergéniques des gènes ribosomiques (ADNr) qui recrutent notamment le complexe de remodelage nucléolaire NoRC et participent à la mise sous silence épigénétique de certaines copies des gènes ribosomiques (contrôle épigénétique de la transcription des ADNr)
- Les ARN associés aux PIWI (piARN ou piwi-interacting RNA) participent principalement au maintien de l'intégrité du génome en réprimant les éléments transposables, notamment dans les cellules germinales, par des mécanismes faisant intervenir la méthylation de l'ADN et les modifications de la chromatine.
- Les microARN (miRNA) agissent essentiellement au niveau post-transcriptionnel en contrôlant la stabilité ou la traduction des ARNm, mais certains contribuent également, directement ou indirectement, à la régulation de mécanismes épigénétiques.
(Figure : vetopsy.fr)
Régulation épigénétique
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation