


Acides nucléiques
ARN : traduction
Initiation
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L'initiation de la traduction assure le recrutement du ribosome sur l'ARN messager et la sélection du codon AUG initiateur grâce à l'action coordonnée des facteurs d'initiation eIF.
Initiation de la traduction
1. L'initiation constitue la première étape de la traduction et correspond à l'ensemble des mécanismes permettant de positionner correctement le ribosome sur l'ARN messager (ARNm) afin de définir le codon d'initiation qui servira de point de départ à la syntdèse protéique.
Chez les eucaryotes, ce processus repose sur l'action coordonnée :
- de la petite sous-unité ribosomique 40S,
- de l'ARN de transfert (ARNt) initiateur chargé de méthionine,
- de nombreux facteurs d'initiation appelés eIF (eukaryotic Initiation Factors),
2 L'initiation conduit successivement :
- à la formation des complexes de pré-initiation 48S,
- au scanning de la région 5'UTR et à la reconnaissance du codon AUG initiateur,
- à l'assemblage du ribosome 80S compétent pour entrer dans la phase d'élongation.
Formation du complexe de pré-initiation 48S
1. L'initiation débute par la préparation de l'ARN messager au niveau de son extrémité 5' (Translation Initiation Regulated by RNA-Binding Protein in Mammals: The Modulation of Translation Initiation Complex by Trans-Acting Factors 2021).
a. La coiffe 7-méthylguanosine (m7G) est reconnue par le facteur eIF4E, une protéine spécialisée dans la liaison à cette structure (Control of the eIF4E activity: structural insights and pharmacological implications 2021).
b. eIF4E recrute ensuite eIF4G, une protéine d'échafaudage capable d'interagir avec de nombreux partenaires impliqués dans l'initiation de la traduction.
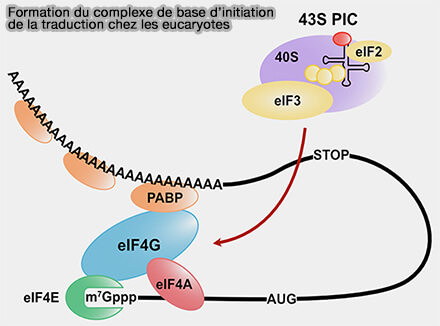
(Figure : vetopsy.fr d'après Fukao et coll)
- Cette interaction repose sur un motif conservé de liaison à eIF4E appelé 4E-BM (eIF4E-binding motif) présent dans eIF4G.
- Certaines protéines régulatrices de eIF4E, notamment les protéines 4E-BP, possèdent également un motif 4E-BM ainsi qu'une région de liaison non canonique supplémentaire (NC:4E-BM, non-canonical eIF4E-binding motif) qui renforce leur affinité pour eIF4E et leur permet d'entrer en compétition avec eIF4G.
c. L'association de eIF4E et eIF4G permet ensuite le recrutement de eIF4A, une hélicase ARN ATP-dépendante appartenant à la famille des protéines DEAD-box, dont l'activité est stimulée par eIF4B, qui participe au déroulement des structures secondaires présentes dans la région 5'UTR de l'ARNm.
L'association de eIF4E, eIF4G et eIF4A forme le complexe eIF4F.
Remarque : les IRES (Internal Ribosome Entry Site) correspondent à des structures d’ARN capables de recruter directement les complexes d’initiation de la traduction indépendamment de la coiffe 5′ habituellement requise pour l’initiation canonique de la traduction (Translation regulation by ribosomes: Increased complexity and expanded scope 2015).
- Ces éléments sont notamment présents dans de nombreux ARN viraux ainsi que dans certains ARNm cellulaires dont la traduction doit être maintenue lorsque l'initiation dépendante de la coiffe est inhibée.
- Les mécanismes moléculaires utilisés par les différents IRES sont très variables et peuvent nécessiter des combinaisons distinctes de facteurs d'initiation ou de protéines auxiliaires spécialisées.
2. Parallèlement, l'extrémité 3' de l'ARNm est reconnue par la protéine PABPC1 (Poly(A)-Binding Protein Cytoplasmic 1), qui se fixe à la queue poly(A) ( protéines PABP).
protéines PABP).
L'interaction entre PABPC1 et eIF4G rapproche les extrémités 5' et 3' de l'ARNm et favorise la formation d'une structure circulaire appelée modèle de boucle fermée (closed-loop model), qui améliore l'efficacité de l'initiation et facilite le recyclage des ribosomes.
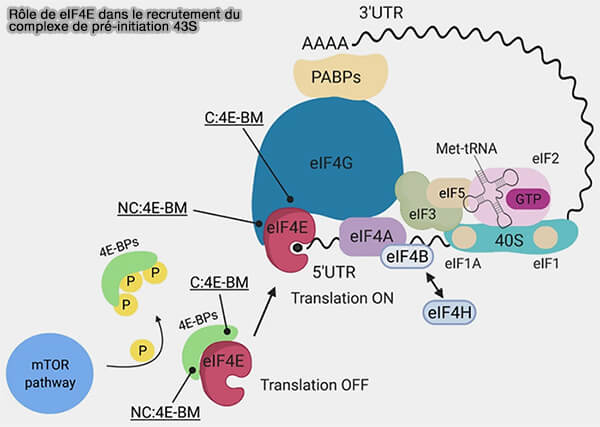
(Figure : vetopsy.fr adaptée d'après Romagnoli et coll)
3. Conjointement à la préparation de l'ARNm, la petite sous-unité ribosomique 40S s'associe à plusieurs facteurs d'initiation pour former le complexe de pré-initiation 43S (43S PIC).
Ce complexe comprend notamment :
- la sous-unité 40S,
- l'ARNt initiateur Met-ARNtiMet,
- eIF2 lié au GTP, qui assure le recrutement de l'ARNt initiateur,
- eIF1 et eIF1A, impliqués dans la sélection correcte du codon d'initiation,
- eIF3, un complexe multiprotéique constitué de plusieurs sous-unités périphériques (eIF3b, eIF3g et eIF3i) organisées autour d'un cœur structural octamérique (eIF3a, eIF3c, eIF3e, eIF3f, eIF3h, eIF3k, eIF3l et eIF3m), qui stabilise le complexe de pré-initiation, favorise son recrutement sur l'ARNm et coordonne plusieurs étapes de l'initiation,
- eIF5, qui participe aux étapes ultérieures de reconnaissance du codon d'initiation.
4. Le complexe 43S est ensuite recruté à l'extrémité 5' de l'ARNm grâce aux interactions entre eIF3 et eIF4G conduisant à la formation du complexe de pré-initiation 48S (The structure of a human translation initiation complex reveals two independent roles for the helicase eIF4A 2024 et Structure of a human 48S translational initiation complex 2021).

(Figure : vetopsy.fr adaptée d'après Brito Querido et coll)
Scanning et reconnaissance du codon d'initiation
Le scanning correspond à la progression du complexe de pré-initiation 48S le long de la région 5'UTR de l'ARNm dans le sens 5' ➞ 3' à la recherche d'un codon d'initiation situé dans un contexte nucléotidique favorable.
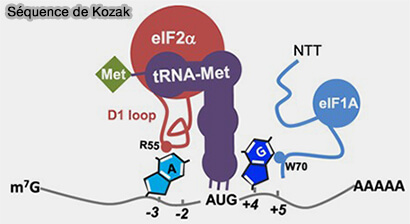
(Figure : vetopsy.fr d'après Hernández et coll)
1. Au cours de ce processus, le complexe de pré-initiation 48S analyse successivement les séquences rencontrées le long de la région 5'UTR afin d'identifier le site de démarrage de la traduction.
a. Lorsque l'anticodon de l'ARNt initiateur Met-ARNtiMet rencontre un codon AUG entouré d'un contexte nucléotidique favorable correspondant à la séquence de Kozak, un appariement stable peut alors s'établir (Functional analysis of the AUG initiator codon context reveals novel conserved sequences that disfavor mRNA translation in eukaryotes 2023).
La séquence de Kozak est caractérisée par la séquence consensus GCC(A/G)CCAUGG, les nucléotides en gras sont les plus importants pour la reconnaissance du codon AUG (Functional analysis of the AUG initiator codon context reveals novel conserved sequences that disfavor mRNA translation in eukaryotes 2023).
b. La reconnaissance de cette séquence favorise l'arrêt du scanning et permet de distinguer le véritable codon d'initiation des autres codons AUG éventuellement présents dans l'ARNm (Selection of start codon during mRNA scanning in eukaryotic translation initiation 2022).
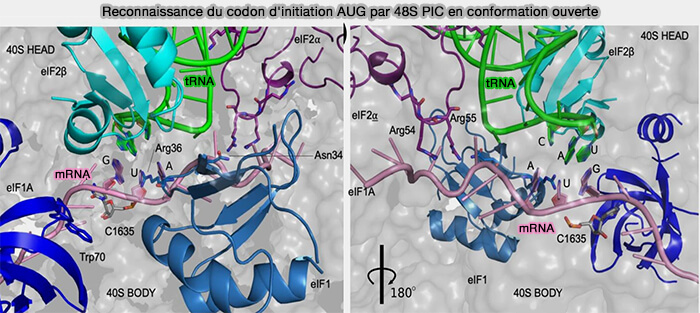
(Figure : vetopsy.fr adaptée d'après Basu et coll)
2. Pendant la phase de scanning, eIF1A contribue, en association avec eIF1, au maintien du complexe de pré-initiation 48S dans une conformation ouverte (" open ") caractérisée par une occupation incomplète du site P de la sous-unité 40S par le Met-ARNtiMet (état POUT).
Dans cette configuration, l'appariement entre l'anticodon de l'ARNt initiateur et les codons successivement rencontrés demeure instable, ce qui permet au complexe de poursuivre sa progression le long de l'ARNm tant qu'un codon AUG situé dans un contexte de Kozak favorable n'a pas été reconnu.
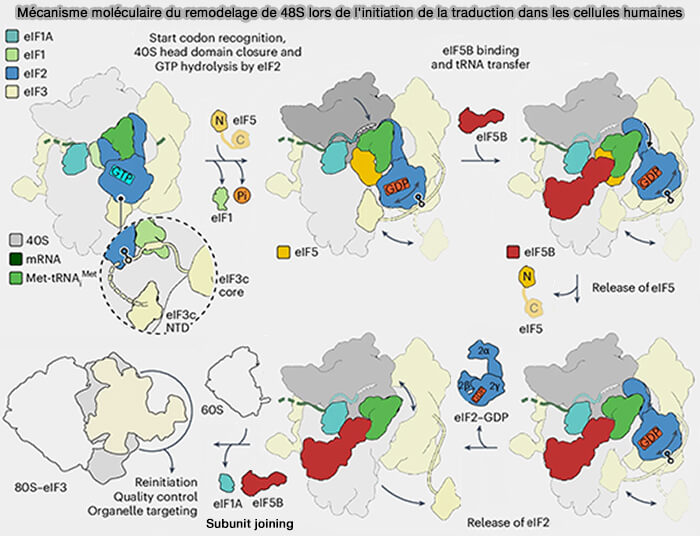
(Figure : vetopsy.fr adaptée d'après Brito Querido et coll)
3. La reconnaissance d'un codon AUG approprié provoque alors une réorganisation du site P et la transition d'un état ouvert (open, POUT) vers un état fermé (closed, PIN), dans lequel le Met-ARNtiMet est pleinement engagé dans le site P (Structural basis for translational control by the human 48S initiation complex 2024).
a. Cet transition :
- entraîne l'arrêt du scanning,
- favorise la dissociation de eIF1,
- permet à eIF5, GAP (GTPase-Activating Protein ou GTPase-Accelerating Protein) de eIF2, de stimuler l'hydrolyse du GTP associé à eIF2 en GDP
b. L'hydrolyse du GTP porté par eIF2 verrouille alors le complexe 48S sur le codon initiateur sélectionné et déclenche les réarrangements conformationnels conduisant à la dissociation de eIF1, de eIF2-GDP et de eIF5, convertissant ainsi le complexe 48S en un complexe prêt à recruter la grande sous-unité ribosomique 60S (Exploring the interaction dynamics of eukaryotic translation initiation factor 2).
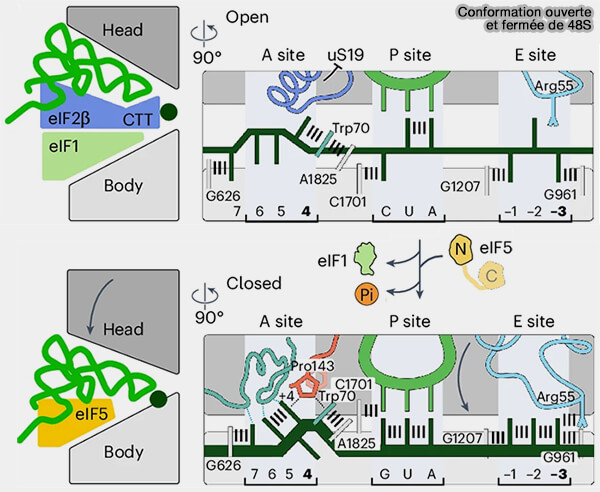
(Figure : vetopsy.fr adaptée d'après Brito Querido et coll)
Assemblage du ribosome 80S
Le complexe 48S validé devient alors compétent pour recruter la grande sous-unité ribosomique 60S.
1. L'association de la sous-unité 60S conduit à la formation d'un complexe 80S transitoire contenant encore eIF5B et à eIF1A.
L'hydrolyse du GTP porté par eIF5B, une GTPase conservée apparentée au facteur bactérien IF2, entraîne alors la dissociation de ces deux derniers facteurs, conduisant à la formation d'un ribosome 80S pleinement fonctionnel (Established and Emerging Regulatory Roles of Eukaryotic Translation Initiation Factor 5B (eIF5B) 2021).
Remarque : les sous-unités 40S et 60S ayant préalablement achevé leur maturation finale dans le cytoplasme, leur association est alors rendue possible lors de l'initiation de la traduction (assemblage des sous-unités pré-ribosomiques).
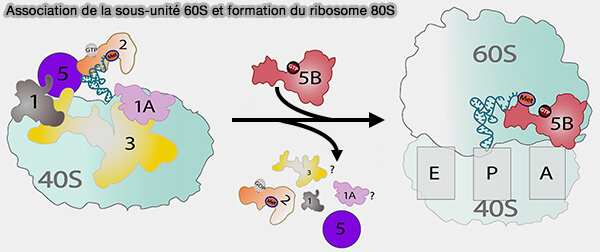
(Figure : vetopsy.fr adaptée d'après Chukka et coll)
2. À l'issue de l'initiation, le Met-ARNtiMet demeure positionné dans le site P du ribosome tandis que le site A reste libre pour accueillir le premier aminoacyl-ARNt de la phase d'élongation.
Le ribosome est alors correctement positionné sur le codon d'initiation et prêt à entrer dans la phase d'élongation de la traduction.
Élongation, terminaison et régulation de la traduction
L'initiation de la traduction se poursuit par plusieurs mécanismes assurant la synthèse, l'arrêt et le contrôle de la production des protéines.
Cette page est consacrée :
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation