


Acides nucléiques
ARN : édition
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Matrice extracellulaire
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L'édition de l'ARN (RNA editing) est un mécanisme post-transcriptionnel qui modifie spécifiquement la séquence nucléotidique de certains ARN par conversion d'une base (A→I, C→U ou, plus rarement, U→C), permettant de produire des molécules différentes de celles directement prédites par la séquence de l'ADN.
Vue d'ensemble
1. Contrairement à la maturation des ARN, qui transforme un transcrit primaire en ARN fonctionnel sans modifier l'information génétique portée par les exons, l'édition de l'ARN modifie directement certains nucléotides après ou, plus rarement, au cours de la transcription.

L'ARN obtenu peut ainsi différer de celui qui serait attendu à partir de la seule séquence de l'ADN.

(Figure : vetopsy.fr d'après Phoebe)
{kind=link}
Le terme " édition " est employé par analogie avec la correction d'un texte avant sa publication : certaines bases sont remplacées, ajoutées ou supprimées de manière ciblée afin de modifier les propriétés de l'ARN.
2. Chez les eucaryotes, l'édition concerne principalement certains :
- ARN messagers (ARNm),
- ARN de transfert (ARNt),
- ARN ribosomiques (ARNr),
- ARN non codants (ncRNA), notamment les microARN (miRNA).
3. Chez les mammifères, elle reste relativement rare, ne touche qu'un nombre limité de transcrits.
a. Elle repose essentiellement sur la désamination enzymatique de certaines bases, conduisant principalement à trois types de modifications :
- la conversion d'une adénosine en inosine (A ➞ I), la plus fréquente chez les animaux et catalysée par les enzymes ADAR ;
- la conversion d'une cytidine en uridine (C ➞ U), catalysée par des enzymes de la famille APOBEC,
- plus rarement, la conversion d'une uridine en cytidine (U ➞ C), observée dans quelques tissus et chez certains organismes.
b. Ces modifications se produisent principalement dans le noyau ou le cytoplasme.
Remarque : un autre mécanisme d'édition est fondé sur l'insertion ou la suppression de nucléotides, et essentiellement observé dans les kinétoplastes des protozoaires des Kinetoplastida,
- Il est observé essentiellement chez les protozoaires de la classe des Kinetoplastida, notamment Trypanosoma brucei.
- Cette édition se déroule dans le kinétoplaste, une région spécialisée de leur unique mitochondrie contenant le kDNA, un réseau de molécules d'ADN circulaires, où elle est assurée par un complexe multiprotéique appelé éditosome, qui catalyse l'insertion et la suppression de nucléotides. (Structure of the mitochondrial editosome-like complex associated TUTase 1 reveals divergent mechanisms of UTP selection and domain organization 2010 et RNA Binding and Core Complexes Constitute the U-Insertion/Deletion Editosome 2014).
Modification d'une base par désamination
Conversion de l'adénosine en inosine (A-to-I editing)
La conversion de l'adénosine en inosine (A ➞ I editing) constitue la forme d'édition de l'ARN la plus fréquente chez les mammifères.

Vous pouvez lire : Structural perspectives on adenosine to inosine RNA editing by ADARs (2024) couvrant l'ensemble des aspects structuraux, moléculaires et fonctionnels de l'édition A ➞ I.
Enzymes ADAR et mécanisme d'édition
La désamination enzymatique d'adénosines présentes principalement dans des régions d'ARN double brin (ARNdb) en inosine réaction est catalysée par les ADAR (Adenosine Deaminases Acting on RNA, EC 3.5.4.37), une famille d'enzymes spécialisée dans l'édition de l'ARN (A-to-I RNA editing - immune protector and transcriptome diversifier 2018).
Ces enzymes sont distinctes de l'adénosine désaminase (ADA), EC 3.5.4.4, impliquée dans la voie de sauvetage indirecte de l'adénosine.
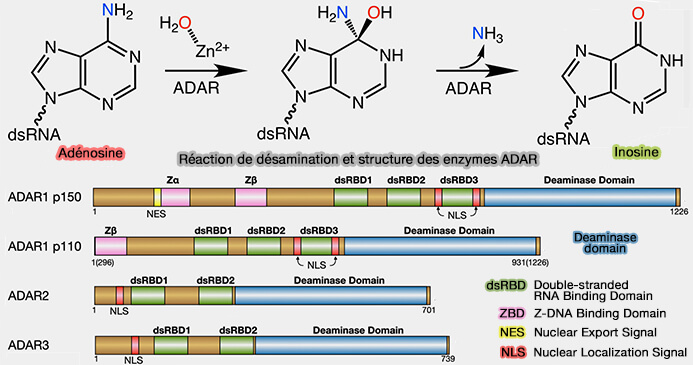
(Figure : vetopsy.fr d'après Fischer et Beal)
1. On distingue trois membres de cette famille chez les mammifères.
a. ADAR1, largement exprimée, existe sous deux isoformes,
- p110, principalement nucléaire,
- p150, induite notamment par les interférons et localisée en grande partie dans le cytoplasme.
b. ADAR2 (ADARB1), largement exprimée, est responsable de la majorité des événements d'édition site-spécifiques des ARNm codants, notamment dans le système nerveux.
c. ADAR3 (ADARB2), exprimée essentiellement dans le cerveau, dépourvue d'activité catalytique, semble agir principalement comme un régulateur négatif de l'activité d'ADAR1 et d'ADAR2, bien que son rôle exact reste débattu.
2. L'inosine n'est pas l'une des quatre bases canoniques de l'ARN, mais est reconnue comme une guanosine par les ribosomes, les ARN polymérases et de nombreuses protéines de liaison aux ARN (RBP).
a. Une conversion A ➞ I est ainsi interprétée fonctionnellement comme une substitution A ➞ G, pouvant modifier la séquence protéique, l'épissage ou la structure secondaire d'un ARN.
- La désamination d'une paire A-U produit une paire I-U, de type wobble, littéralement " appariement bancal ", moins stable que la paire d'origine,.
- La conversion d'un mésappariement A-C génère une paire I-C, plus stable.
b. Ces modifications influencent le repliement des ARN ainsi que leurs interactions avec de nombreuses protéines.
Édition spécifique et hyperédition
On distingue deux principaux modes d'édition qui diffèrent par le nombre d'adénosines modifiées et par leurs conséquences biologiques (A-to-I RNA editing and hematopoiesis 2024).
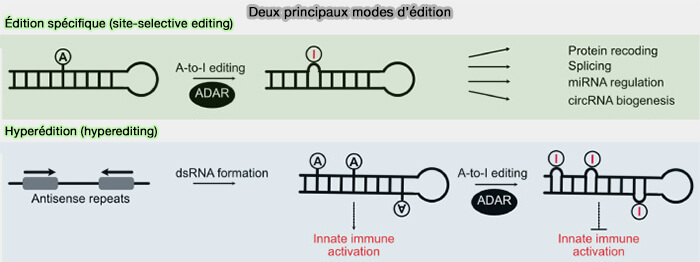
(Figure : vetopsy.fr d'après Liang et coll)
1. L'édition spécifique (site-selective editing) concerne une ou quelques adénosines d'un transcrit donné et touche le plus souvent les régions codantes.
En modifiant un codon, un site d'épissage ou une région régulatrice, elle peut altérer la séquence d'une protéine, modifier l'épissage alternatif ou influencer la stabilité, la localisation et la traduction d'un ARN.
2. L'hyperédition (hyperediting) convertit un grand nombre d'adénosines d'une même région d'ARN double brin et concerne surtout les régions non codantes, riches en répétitions Alu.
Ce mécanisme modifie profondément la structure secondaire de l'ARN et ses interactions avec de nombreuses protéines, influençant ainsi sa stabilité, sa localisation, son épissage, sa traduction, sa dégradation ainsi que la maturation de certains miARN.
3. L'édition A→I peut concerner la plupart des catégories d'ARN cellulaires (Advances in Detection Methods for A-to-I RNA Editing 2025).
a. Chez les mammifères, elle affecte :
- principalement les pré-ARNm, où elle peut modifier les régions codantes, les introns ou les régions non traduites (UTR),
- les miARN, les lncARN et les ARN circulaires (circARN).
b. Selon le type d'ARN concerné et le site édité, elle peut modifier :
- la séquence d'une protéine,
- l'épissage alternatif,
- la maturation des miARN,
- la stabilité ou la localisation des transcrits,
- les interactions entre les ARN et leurs partenaires protéiques.
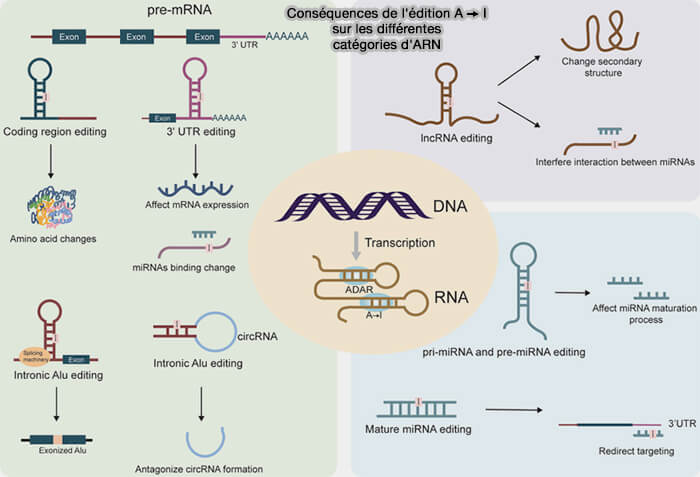
(Figure : vetopsy.fr d'après Yang et Sakurai)
Importance physiologique et pathologique
1. Dans le système nerveux central, l'édition A ➞ I est particulièrement abondante.
a. Elle module l'excitabilité neuronale et la neurotransmission. notamment dans la régulation (RNA Editing and Modifications in Mood Disorders 2020) :
- des récepteurs AMPA du glutamate, en particulier la sous-unité GluA2 (GRIA2)
- du récepteur sérotoninergique 5-HT2C,
- de certains canaux potassiques voltage-dépendants, notamment Kv1.1 (KCNA1).
b. Par exemple, le pré-ARNm du récepteur 5-HT2C est édité sur cinq sites (A à E) situés dans l'exon V, générant plusieurs isoformes présentant des propriétés fonctionnelles différentes et modulant la transmission sérotoninergique.
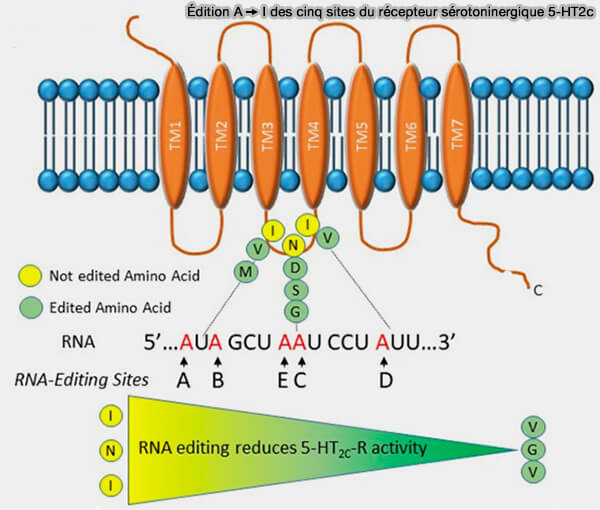
(Figure : vetopsy.fr d'après Barbon et Magri)
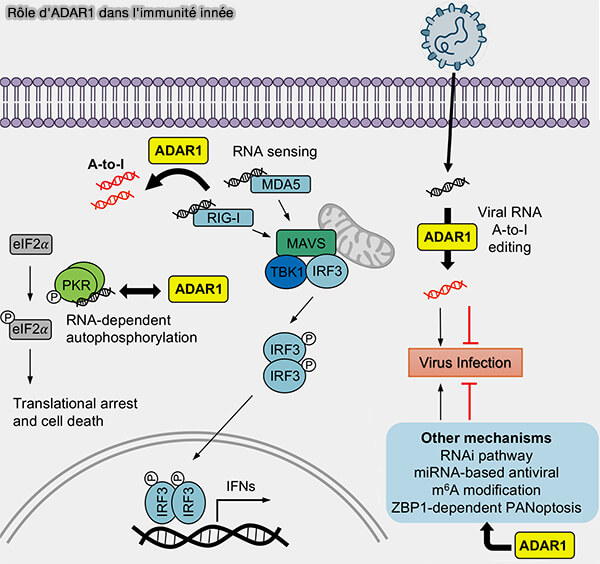
2. Chez les mammifères, ADAR1 joue également un rôle essentiel dans l'immunité innée.
Les cellules produisent naturellement certains ARN double brin endogènes, proviennent notamment chez l'homme de l'appariement de séquences répétées Alu transcrites en orientation inverse au sein d'un même ARN ( SINE ou Short Interspersed Nuclear Elements).
SINE ou Short Interspersed Nuclear Elements).
a. En convertissant des adénosines en inosines dans ces ARN, ADAR1 modifie leurs propriétés et empêche leur reconnaissance par les récepteurs cytoplasmiques de l'immunité antivirale, notamment MDA5 (Melanoma Differentiation-Associated protein 5) et et, dans une moindre mesure, RIG-I (ADAR1: a central regulator of dsRNA sensing in host-virus interactions 2026).
b. Cette édition empêche l'activation de la protéine adaptatrice mitochondriale MAVS (Mitochondrial Antiviral Signaling protein), indispensable à la transmission du signal antiviral.
Elle évite ainsi l'activation des facteurs de transcription IRF3 (Interferon Regulatory Factor 3), responsables de l'expression des interférons de type I, et prévient une réponse immunitaire dirigée contre les ARN de la cellule elle-même, contribuant ainsi au maintien de la tolérance au soi.
c. Cette édition évite ainsi une activation inappropriée de la voie des interférons de type I contre les ARN de la cellule elle-même et contribue au maintien de la tolérance au soi.
(Figure : vetopsy.fr d'après Barbon et Magri)
3. L'altération de l'édition A ➞ I peut entraîner des conséquences pathologiques importantes.
- Des anomalies de l'édition A → I ont également été mises en évidence dans plusieurs maladies neurologiques et neuropsychiatriques, où elles perturbent notamment l'édition de récepteurs et de canaux ioniques impliqués dans la neurotransmission (Spatiotemporal and genetic regulation of A-to-I editing throughout human brain development 2022 et RNA Editing Signatures Powered by Artificial Intelligence: A New Frontier in Differentiating Schizophrenia, Bipolar, and Schizoaffective Disorders 2024).
- Chez l'Homme, des mutations du gène ADAR1 sont responsables du " syndrome d'Aicardi-Goutières ", une maladie génétique rare caractérisée par une activation chronique de la réponse aux interférons de type I, secondaire à une reconnaissance anormale des ARN cellulaires comme des ARN viraux (A-to-I RNA editing by ADAR and its therapeutic applications: From viral infections to cancer immunotherapy 2023).
- Enfin, une dérégulation de l'activité des enzymes ADAR, en particulier ADAR1 et ADAR2, est observée dans de nombreux cancers (The role of ADAR1 through and beyond its editing activity in cancer 2024).
Selon le contexte cellulaire, cette dérégulation peut favoriser ou, au contraire, limiter la progression tumorale en modifiant l'expression de gènes impliqués dans la prolifération, la différenciation, l'apoptose, les interactions avec le microenvironnement tumoral ou encore les réponses immunitaires antitumorales
Conversion de la cytidine en uridine (C-to-U editing)
1. La conversion de la cytidine en uridine (C ➞ U editing) est beaucoup plus rare que l'édition A ➞ I chez les mammifères.
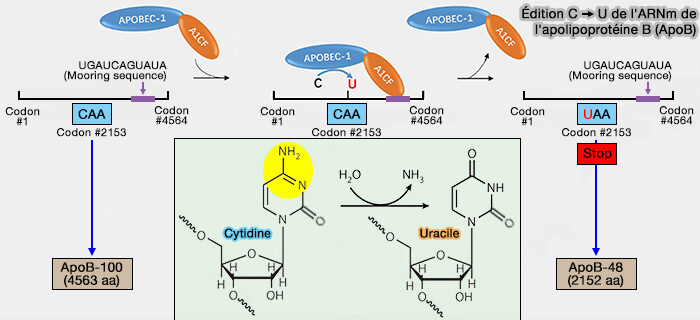
- Elle résulte de la désamination enzymatique, principalement catalysée par APOBEC1 (Apolipoprotein B mRNA Editing Catalytic Polypeptide 1), une cytidine désaminase appartenant à la famille des enzymes APOBEC (APOBEC-1 Complementation Factor: From RNA Binding to Cancer 2024).
- Le mécanisme débute par la reconnaissance de la séquence d'ancrage (mooring sequence), une courte séquence nucléotidique située en aval du site d'édition, par le facteur de complémentation A1CF (APOBEC1 Complementation Factor).
- A1CF recrute ensuite et positionne APOBEC1 sur la cytidine cible, permettant sa désamination en uridine.
(Figure : vetopsy.fr d'après Wang et Cheng)
2. Le principal exemple concerne l'ARNm de l'apolipoprotéine B (ApoB).
a. Dans le foie, l'ARNm d'APOB est traduit en apoB-100 l'apoB-100, ou apoB pleine longueur, i.e. 4563 résidus, car Apobec-1 n'est pratiquement pas exprimée dans le foie humain (apoB-100 et apoB-48).
L'apoB-100 est indispensable à l'assemblage et à la sécrétion des VLDL (lipoprotéines de très basse densité).
b. Dans l'intestin, APOBEC1 convertit le codon CAA en UAA, créant un codon stop prématuré (ApoB48 as an Efficient Regulator of Intestinal Lipid Transport 2020).
Cette édition interrompt la traduction et produit ApoB-48, une protéine de 2 152 acides aminés.
- L'apoB-48 est la principale apolipoprotéine structurale des chylomicrons (CM) et de ses remnants (CM-R) et ne possède pas le domaine C-terminal de liaison au récepteur des LDL (LDLR ou Low-Density Lipoprotein Receptor), ce qui explique ses propriétés métaboliques différentes de celles de l'apoB-100.
Contrairement à A ➞ I, ici l'édition ne sert pas principalement à moduler des ARN, mais à produire deux protéines distinctes à partir d'un même gène.
Conversion de l'uridine en cytidine (U-to-C editing)
La conversion de l'uridine en cytidine (U ➞ C editing) a été décrite dans quelques ARN de mammifères, notamment dans certains transcrits exprimés dans le cerveau, le rein ou le côlon.
Toutefois, contrairement aux éditions A → I et C → U, ce mécanisme reste peu fréquent et son enzyme responsable n'a pas encore été clairement identifiée.
Quelques cas ont notamment été rapportés pour le gène TPH2 (Thrytophane Hydroxylase 2), impliqué dans la biosynthèse de la sérotonine, où cette édition pourrait moduler l'activité de l'enzyme.
Ces observations restent toutefois débattues et nécessitent encore des confirmations indépendantes).
Remarque : chez les plantes, les éditions C → U et U → C sont beaucoup plus fréquentes et interviennent principalement dans les ARN des mitochondries et des chloroplastes (C-to-U and U-to-C: RNA editing in plant organelles and beyond 2023).
Épitranscriptome et autres mécanismes de
régulation post-transcriptionnelle
L'édition de l'ARN agit en complément de plusieurs mécanismes de régulation post-transcriptionnelle des ARN qui contrôlent le devenir et l'expression des transcrits sans modifier la séquence de l'ADN, notamment :
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation