


Acides nucléiques
ARN : dégradation
Dégradation cytoplasmique des ARNm
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La dégradation cytoplasmique des ARN messagers repose sur des mécanismes de déadénylation, de décapping et de surveillance qui contrôlent la stabilité, la traduction et le devenir des ARNm.
Stabilité et durée de vie des ARNm
Les ARN messagers (ARNm) présents dans le cytoplasme possèdent une durée de vie variable qui dépend de mécanismes contrôlant leur stabilité, leur traduction et leur dégradation.
1. Chez les eucaryotes, la stabilité des ARNm constitue un paramètre majeur de régulation de l’expression génique, car elle détermine la durée pendant laquelle un transcrit peut être traduit en protéine ( traduction).
traduction).
- La demi-vie des ARN cytoplasmiques varie fortement selon les transcrits et les conditions cellulaires.
- Certains ARN messagers très instables ne persistent que quelques minutes, tandis que d’autres peuvent rester présents pendant plusieurs heures voire plusieurs jours dans le cytoplasme.
2. Cette stabilité dépend notamment :
- de la structure de la coiffe en 5′,
- de la longueur de la queue poly(A),
- de la présence de séquences régulatrices dans les régions non traduites (UTR),
- de l’association des ARN à des protéines de liaison aux ARN (RBP),
- de leur état de traduction par les ribosomes,
- de l’action d'ARN non codants régulateurs comme les microARN (miRNA).
3. Les régions non traduites des ARN messagers jouent un rôle central dans le contrôle de leur stabilité et certaines séquences présentes dans les régions 3′UTR ou 5′UTR peuvent favoriser :
- la stabilisation des ARN,
- leur dégradation rapide,
- leur stockage transitoire,
- leur adressage vers des mécanismes spécifiques de surveillance cytoplasmique.

Les mécanismes spécifiques de surveillance cytoplasmique des ARNm, étroitement couplés à la traduction et à la progression des ribosomes, sont étudiés dans un chapitre spécifique.
4. La dégradation des ARNm cytoplasmiques repose principalement sur deux grandes catégories de mécanismes :
- les voies canoniques de renouvellement des ARNm,
- les voies de dégradation dirigée impliquant notamment les ARN non codants régulateurs.
5. La traduction et la dégradation des ARN cytoplasmiques sont étroitement couplées et et au devenir des ARN messagers dans le cytoplasme (devenir des ARN messagers cytoplasmiques).
Dégradation canonique des ARN messagers
La dégradation des ARN messagers cytoplasmiques repose principalement sur deux grandes voies exoribonucléolytiques qui impliquent soit une dégradation dans le sens 5′ ➞ 3′, soit une dégradation dans le sens 3′ ➞ 5′.
Chez les eucaryotes, ces mécanismes sont étroitement couplés à des modifications progressives des extrémités des ARN messagers, et la dégradation des ARNm suit généralement une série d’étapes successives (Rôle de la protéine EB2 du virus d’Epstein-Barr dans le métabolisme des ARN messagers 2016).
Déadénylation de la queue poly(A)
La première étape correspond à un raccourcissement progressif de la queue poly(A) appelé déadénylation.
- La queue poly(A), associée aux protéines PABP (poly(A)-binding proteins), stabilise les ARN messagers et favorise leur traduction en facilitant les interactions entre les protéines liées à la queue poly(A) et les complexes d’initiation associés à la coiffe 5′ des transcrits.
- Le raccourcissement progressif de cette queue diminue la fixation des protéines PABP, déstabilise l’organisation circulaire fonctionnelle de l’ARNm et favorise son engagement dans les voies de dégradation cytoplasmique.
Chez les eucaryotes, la déadénylation est principalement assurée par deux grands complexes multiprotéiques.
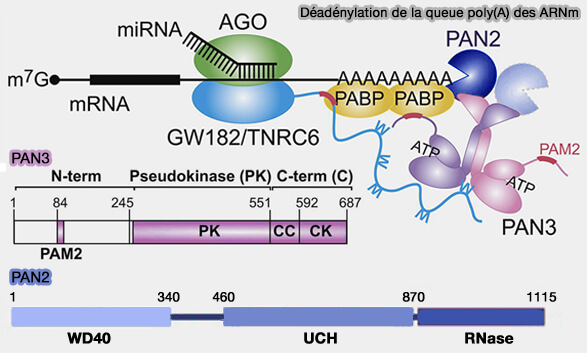
(Figure : vetopsy.fr adaptée d'après Christie et coll)
1. Le complexe PAN2-PAN3 (Poly(A) Nuclease) intervient généralement dans les premières étapes du raccourcissement de la queue poly(A), en particulier lorsque celle-ci reste encore fortement associée aux protéines PABP (Structure of the PAN3 Pseudokinase Reveals the Basis for Interactions with the PAN2 Deadenylase and the GW182 Proteins 2013 et mRNA Deadenylation by Pan2/Pan3 2014).
- PAN2 correspond à la sous-unité catalytique possédant l’activité exoribonucléase 3′ → 5′.
- PAN3, organisée sous forme dimérique, agit principalement comme sous-unité régulatrice capable d’interagir, via son domaine PAM2 (PABP-interacting motif 2), avec les protéines PABP associées à la queue poly(A), favorisant ainsi le recrutement et l’activité du complexe sur les ARN messagers polyadénylés.
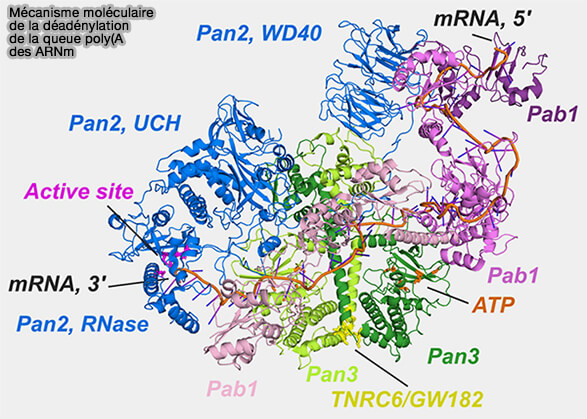
(Figure : vetopsy.fr adaptée d'après Krempl et coll)
2. Le complexe CCR4-NOT (Carbon Catabolite Repression 4-CCR4-NOT transcription complex) constitue le principal complexe de déadénylation cytoplasmique chez les eucaryotes et et assure l’essentiel du raccourcissement de la queue poly(A) (A structural biology view on the enzymes involved in eukaryotic mRNA turnover 2023).
a. Ce complexe multiprotéique comprend notamment (Regulation of the multisubunit CCR4-NOT deadenylase in the initiation of mRNA degradation 2023) :
- la grande sous-unité NOT1, qui forme la plateforme centrale d’assemblage du complexe CCR4-NOT,
- les sous-unités NOT2 et NOT3, qui stabilisent l’organisation du complexe et participent à son architecture globale,
- le module catalytique CAF1-CCR4, le module catalytique CAF1-CCR4, comprenant notamment les nucléases CCR4a/CNOT6, CCR4b/CNOT6L, CAF1/CNOT7-CNOT8, responsables de l’activité de déadénylation exoribonucléolytique 3′ → 5′,
- le module CAF40, impliqué dans les interactions avec différents facteurs régulateurs et dans le recrutement du complexe vers certains ARN messagers cibles,
- le module NOT10-NOT11, qui participe à la stabilité structurale du complexe CCR4-NOT et à son organisation fonctionnelle dans le cytoplasme.
Remarque : dans la nomenclature humaine, les protéines NOT sont souvent désignées CNOT, par exemple NOT1/CNOT1…
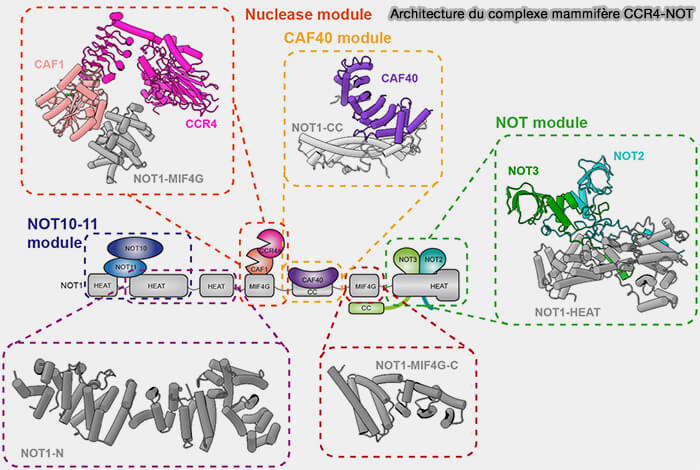
(Figure : vetopsy.fr adaptée d'après Raisch et Valkov)
b. Le complexe CCR4-NOT peut être recruté vers les ARN messagers par différents mécanismes, notamment (Regulation of the multisubunit CCR4-NOT deadenylase in the initiation of mRNA degradation 2023 qui décrit les différentes interactions) :
- des protéines de liaison aux ARN (RBP) capables de reconnaître des séquences ou structures spécifiques présentes dans les ARN messagers et de recruter le complexe CCR4-NOT vers les transcrits cibles,
- les protéines GW182/TNRC6 associées aux complexes miRNA-Argonaute,
- certains mécanismes de surveillance traductionnelle des ARN messagers, notamment la voie NMD (nonsense-mediated decay), capables de coupler la reconnaissance des transcrits aberrants au recrutement des mécanismes de déadénylation.
3. Après raccourcissement important de la queue poly(A), les ARN messagers deviennent plus accessibles aux mécanismes de décapping et/ou aux exoribonucléases cytoplasmiques impliquées dans leur dégradation complète.
Remarque : certaines déadénylases spécialisées, comme PARN (Poly(A)-specific ribonuclease), interviennent dans la maturation ou la surveillance de certains ARN cellulaires, notamment des ARN non codants et de l’ARN de la télomérase.
Dégradation 5′ ➞ 3′ et 3′ ➞ 5′
Après déadénylation, les ARN messagers peuvent être orientés vers deux grandes voies de dégradation cytoplasmique :
- une voie assurée par des exoribonucléases 5′ → 3′.
- une voie assurée par des exoribonucléases 5′ → 3′.
Dégradation 5′ ➞ 3′
Dans la voie de dégradation 5′ ➞ 3′, les ARN messagers subissent tout d'abord un décapping correspondant à l’élimination de la coiffe m7G et l'exposition de l’extrémité 5′ du transcrit aux exoribonucléases cytoplasmiques (mRNA decapping: finding the right structures 2018).
Décapping des ARN messagers
1. Le décapping (décoiffage) est principalement assuré par le complexe DCP1-DCP2 qui en constitue le cœur.
- DCP2 correspond à la sous-unité catalytique qui hydrolyse la coiffe 5′ des ARN messagers et génère une extrémité 5′ monophosphate reconnue par l’exoribonucléase XRN1.
- DCP1 agit principalement comme un facteur activateur et régulateur de DCP2, en favorisant son activité de décapping.
2. Plusieurs cofacteurs contribuent au recrutement et à l’activation du complexe DCP1-DCP2.
a. EDC3 (Enhancer of mRNA Decapping 3), protéine activatrice du décapping, est capable de dimériser et d’interagir avec plusieurs protéines impliquées dans la répression traductionnelle et la dégradation des ARN messagers.
b. L’hélicase DDX6 interagit notamment avec :
- PATL1 (Protein Associated with Topoisomerase II Homolog 1), une protéine adaptatrice impliquée dans la répression traductionnelle et le recrutement des facteurs de décapage,
- le complexe CCR4-NOT, contribuant ainsi au couplage fonctionnel entre déadénylation, répression traductionnelle et dégradation 5′ → 3′ des ARNm
DDX6 agit comme une ATPase de remodelage des complexes ribonucléoprotéiques associés aux ARN messagers (mRNP), favorisant la dissociation des facteurs de traduction, la transition des ARNm vers un état réprimé et l’exposition de la coiffe au complexe DCP1-DCP2.
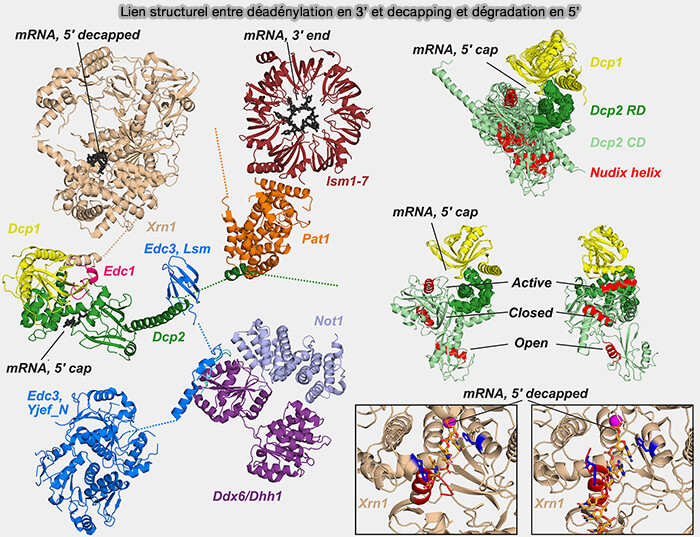
(Figure : vetopsy.fr adaptée d'après Krempl et coll)
c. Le complexe LSM1-7 se fixe préférentiellement aux extrémités 3′ oligoadénylées ou fortement raccourcies des ARNm après déadénylation et favorise le recrutement du complexe de décapping via la protéine PATL1, elle-même associée à LSM1-7 et reliée à DCP2.
Le complexe cytoplasmique LSM1-7 impliqué dans le décapping et la dégradation des ARN messagers est distinct du complexe nucléaire LSM2-8 associé au snARN U6 et au fonctionnement du spliceosome ( protéines LSm)
Remarque : chez la levure, Edc1 se fixe au complexe Dcp1-Dcp2 et favorise l’alignement du domaine catalytique Nudix de Dcp2 avec le substrat coiffé, stabilisant ainsi la conformation active nécessaire à l’hydrolyse de la coiffe m7G.
Chez les métazoaires, le complexe de décapping est également organisé autour d’EDC4 (Ge-1/Hedls), une protéine échafaudage assurant la stabilisation et l’assemblage de multiples facteurs du décapping, notamment DCP1, DCP2, XRN1 et plusieurs cofacteurs régulateurs associés aux P-bodies cytoplasmiques (The activation of the decapping enzyme DCP2 by DCP1 occurs on the EDC4 scaffold and involves a conserved loop in DCP1 2014).
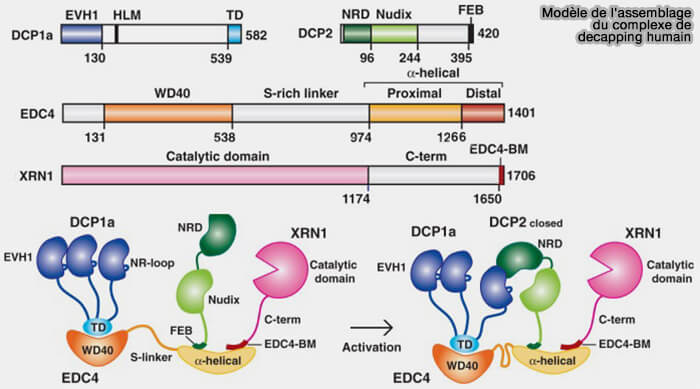
(Figure : vetopsy.fr adaptée d'après Chang et coll)
Dégradation 5′ ➞ 3′ par XRN1
Les ARN décapés sont ensuite rapidement dégradés dans le cytoplasme par l’exoribonucléase XRN1, enzyme hautement processive agissant dans le sens 5′ ➞ 3′ et participant ainsi au renouvellement basal des ARN messagers ainsi qu’à l’élimination de nombreux transcrits ciblés par des mécanismes de régulation post-transcriptionnelle ou de surveillance des ARN (XRN 5’→3’ exoribonucleases: Structure, mechanisms and functions 2014).
1. XRN1 reconnaît préférentiellement les ARN portant une extrémité 5′ monophosphate générée après l’élimination de la coiffe par le complexe DCP1-DCP2.
- Cette spécificité assure un couplage étroit entre le décapping et la dégradation 5′ → 3′ des ARN messagers.
- Après fixation à l’extrémité 5′ du transcrit, XRN1 dégrade l’ARN de manière processive sans se dissocier après chaque événement catalytique, ce qui permet une élimination rapide et efficace des ARN messagers décapés dans le cytoplasme.
2. XRN1 est fréquemment associée aux P-bodies, condensats cytoplasmiques enrichis en facteurs impliqués dans la répression traductionnelle, le décapping et la dégradation des ARN messagers, mais peut également agir dans le cytoplasme diffus.
Remarque : chez les eucaryotes, XRN2 est principalement localisée dans le noyau et participe notamment à certains mécanismes de terminaison transcriptionnelle après le clivage des pré-ARNm (exoribonucléases XRN).
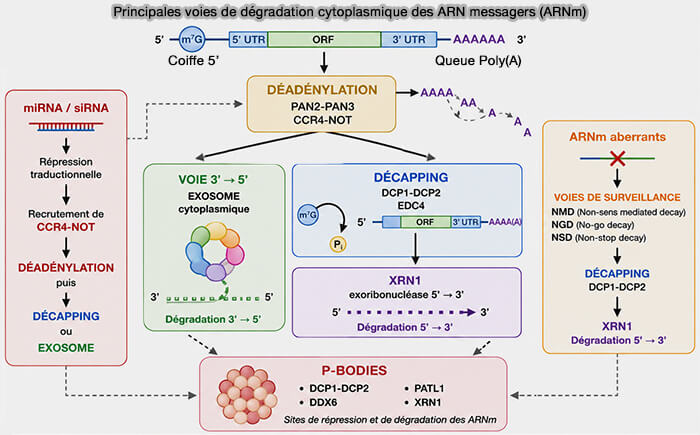
(Figure : vetopsy.fr)
Dégradation 3′ ➞5′
Dans la voie de dégradation 3′ ➞ 5′, les ARN messagers sont d’abord soumis à un raccourcissement important de leur queue poly(A), ce qui favorise leur prise en charge par le complexe de l’exosome associé au complexe SKI.
- Dans cette voie, les ARN sont progressivement dégradés à partir de leur extrémité 3′ sans nécessiter obligatoirement d’étape initiale de décapping, la coiffe pouvant être éliminée ultérieurement lorsque le transcrit devient fortement raccourci ou partiellement dégradé.
L'exosome et le complexe SKI sont étudiés dans un chapitre spécifique.
Dans les deux voies, la dégradation progressive des ARN messagers conduit finalement à la production de petits fragments d’ARN puis de nucléotides monophosphates recyclés par le métabolisme cellulaire.
Dégradation dirigée par les ARN régulateurs
Chez les eucaryotes, certains petits ARN régulateurs peuvent orienter spécifiquement des ARN cibles vers des mécanismes de répression traductionnelle et de dégradation cytoplasmique.
Ces mécanismes participent au contrôle post-transcriptionnel de l’expression génique et permettent une régulation fine, rapide et spécifique de nombreux ARN cellulaires.
Les principaux ARN impliqués dans ces processus sont les microARN (miRNA) et les petits ARN interférents (siRNA) associés aux protéines Argonaute au sein du complexe RISC (RNA-induced silencing complex).
Le mécanisme d'action détaillé des miRNA est étudié dans un chapitre spécifique.
1. Les miRNA induisent principalement une répression traductionnelle associée à une déstabilisation progressive des ARN messagers cibles ( dégradation par les miRNA).
- Chez les animaux, l’appariement entre miRNA et ARN cible est généralement partiel et conduit au recrutement de complexes de déadénylation comme CCR4-NOT, au raccourcissement progressif de la queue poly(A), au décapping des ARN messagers, puis à leur dégradation par XRN1 ou par l’exosome cytoplasmique.
- Lorsque la complémentarité entre le petit ARN guide et l’ARN cible est très élevée, certaines protéines Argonaute possédant une activité endoribonucléase dite " slicer " peuvent cliver directement l’ARN cible.
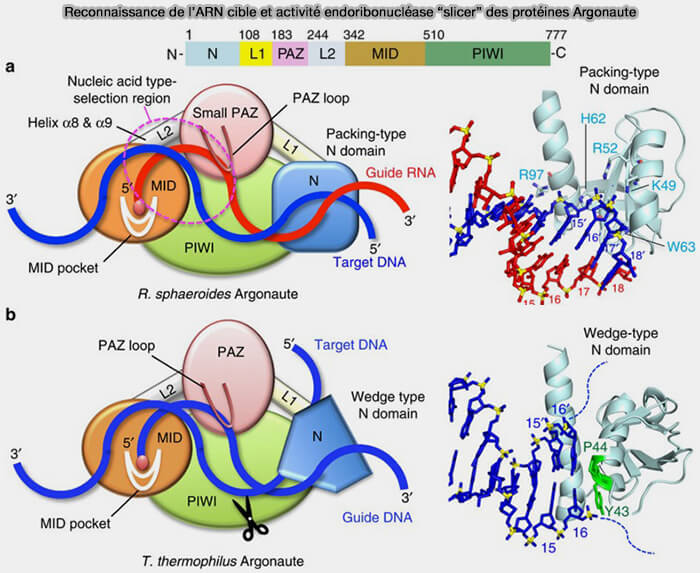
(Figure : vetopsy.fr d'après Miyoshi et coll)
2. Les siRNA induisent généralement une dégradation plus directe des ARN cibles car la complémentarité entre le petit ARN guide et l’ARN cible est quasi parfaite.
Le clivage direct de l’ARN cible par Argonaute génère alors des fragments rapidement dégradés par les systèmes cytoplasmiques de dégradation des ARN, notamment par l’exoribonucléase XRN1 et par l’exosome cytoplasmique associé au complexe SKI.
3. La dégradation dirigée par les ARN régulateurs coopère étroitement avec les mécanismes généraux de surveillance et de dégradation cytoplasmique des ARN messagers.
Devenir cytoplasmique des ARN messagers (ARNm)
La dégradation ne constitue qu’un des devenirs possibles des ARN messagers dans le cytoplasme.
Selon leur état fonctionnel et les signaux cellulaires reçus, les ARNm peuvent être orientés vers la traduction, le stockage, la répression traductionnelle ou la dégradation.
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation