Les mécanismes de surveillance des ARN assurent le contrôle qualité des ARN cellulaires en détectant et en éliminant les ARN aberrants produits lors de la transcription, de la maturation ou de la traduction.
Les mécanismes de surveillance des ARN sont essentiels pour :
limiter l’accumulation d’ARN potentiellement délétères susceptibles de conduire à la synthèse de protéines tronquées, mal repliées ou toxiques,
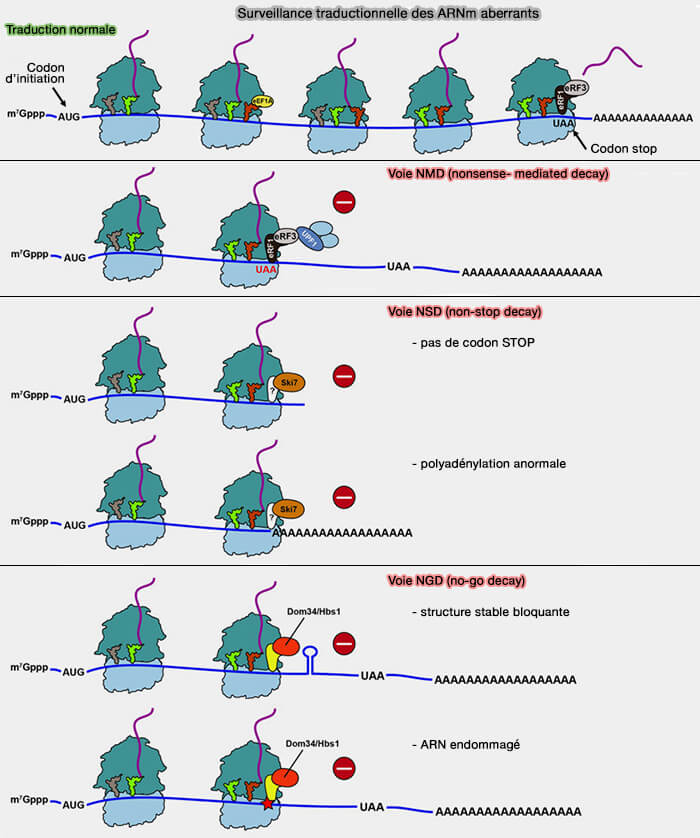
La surveillance traductionnelle repose principalement sur la détection d’anomalies affectant la progression, la terminaison ou le recyclage des ribosomes au cours de la traduction afin de limiter la production de protéines tronquées, aberrantes ou potentiellement toxiques pour la cellule, notamment :
associées aux facteurs de terminaison eRF1 (eukaryotic Release Factor 1) et eRF3.
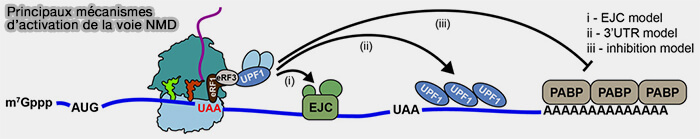
2. Dans la majorité des cas, cette reconnaissance est liée à la persistance d’un complexe EJC (exon junction complex) en aval du codon stop prématuré après l’épissage.
Mécanismes d'activation de la voie NMD
(Figure : vetopsy.fr d'après Simms et coll)
Certaines voies de NMD peuvent également être activées indépendamment des complexes EJC notamment lorsque :
le codon stop est situé à une distance anormalement importante de la queue poly(A), l’éloignement des protéines PABP (poly(A)-binding proteins) associées à la queue poly(A) favorisant alors la stabilisation de UPF1 et l’activation de la voie NMD malgré l’absence de complexes EJC persistants en aval du codon stop,
l'ARNm possède une région 3′UTR particulièrement longue, créant un contexte de terminaison de la traduction anormalement éloigné des facteurs associés à l’extrémité 3′ de l’ARNm.
3. Lors de la première traduction de l’ARNm, les ribosomes déplacent normalement les complexes EJC au cours de leur progression.
Dans un ARNm normal, le ribosome atteint le codon stop physiologique après avoir éliminé la majorité des complexes EJC présents sur l’ARNm.
Au niveau du codon stop, les facteurs de terminaison eRF1 et eRF3 induisent alors la terminaison normale de la traduction et la libération de la chaîne polypeptidique.
La protéine UPF1 peut être recrutée transitoirement au niveau du ribosome en terminaison, mais en l’absence de complexes EJC persistants en aval et donc d’interactions avec UPF2 et UPF3, la voie NMD n’est pas activée.
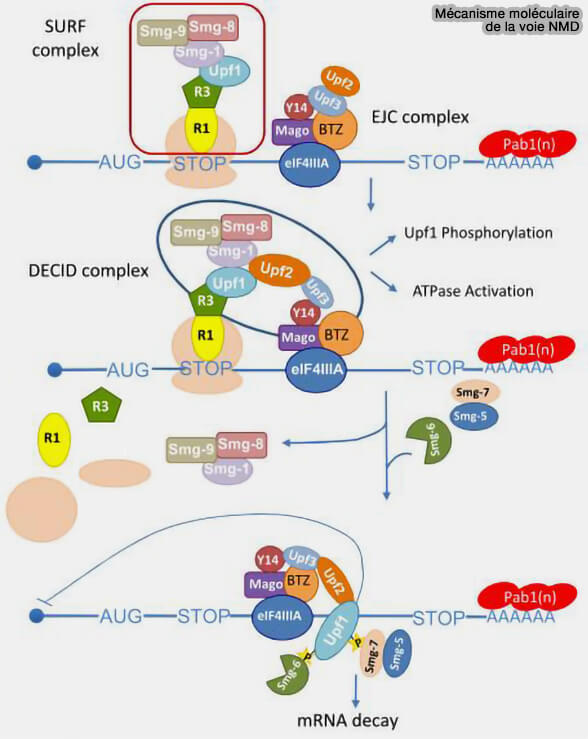
a. Au niveau de ce codon stop prématuré, les facteurs de terminaison eRF1 et eRF3 induisent malgré tout la terminaison de la traduction et recrutent notamment UPF1 ainsi que la kinase SMG1 au voisinage du ribosome arrêté, formant un complexe précoce de surveillance appelé complexe SURF (SMG1-UPF1-eRF1-eRF3).
b. Cependant, contrairement à une terminaison normale, plusieurs complexes EJC persistent encore en aval du ribosome, ce qui montre que la traduction s’est arrêtée prématurément avant la fin normale du cadre de lecture (ORF).
Les protéines UPF3 associées aux complexes EJC persistants en aval peuvent alors interagir, via UPF2, avec UPF1 recrutée au niveau du complexe de terminaison ribosomique.
Au niveau du codon stop prématuré, UPF1, eRF1 et eRF3 (R1 et R3 sur la figure) et la kinase SMG1 associée aux protéines SMG8 et SMG9 forment un complexe de surveillance appelé complexe SURF (SMG1-UPF1-eRF1-eRF3).
Les interactions entre UPF2-UPF3 associés aux complexes EJC persistants et le complexe SURF favorisent alors le rapprochement de ces différents complexes sur l’ARNm et induisent l’activation de UPF1 par phosphorylation via SMG1, conduisant à la formation d’un complexe activateur du NMD parfois désigné complexe DECID (decay-inducing complex).
Mécanisme moléculaire de la voie NMD
(Figure : vetopsy.fr d'après Keverstin et Jacobson)
c. La phosphorylation de UPF1 par SMG1 crée une plateforme de recrutement pour plusieurs protéines régulatrices du NMD, notamment SMG5, SMG6 et SMG7, qui orientent ensuite l’ARNm aberrant vers les mécanismes de dégradation.
SMG6 possède une activité endonucléolytique capable de cliver directement l’ARNm à proximité du codon stop prématuré.
Les complexes SMG5-SMG7 favorisent principalement le recrutement des mécanismes de déadénylation, de décapping et de dégradation exoribonucléolytique des ARN cibles (dégradation canonique des ARN messagers).
Les protéines SMG5 et SMG7 participent également au recyclage du complexe NMD en favorisant la déphosphorylation de UPF1 après l’engagement de la dégradation de l’ARNm cible.
Voie NSD (non-stop decay)
Reconnaissance des ARN messagers dépourvus de codon stop
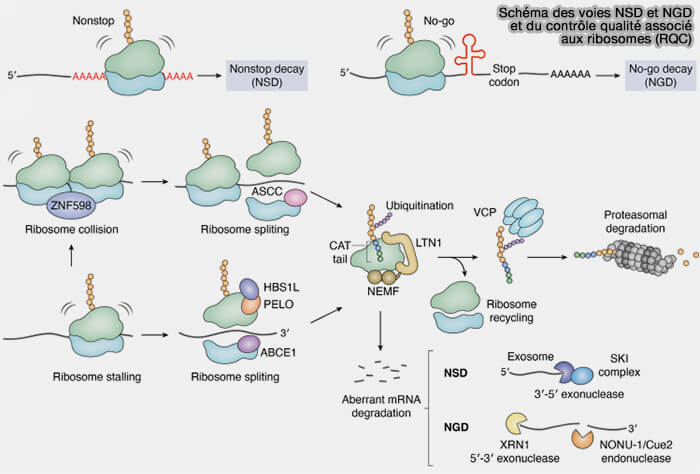
1. La voie NSD (non-stop decay) cible les ARN messagers dépourvus de codon stop fonctionnel.
de mutations supprimant le codon stop physiologique.
b. Dans cette situation, le ribosome ne peut pas recruter correctement les facteurs de terminaison eRF1 et eRF3 normalement associés à la reconnaissance des codons stop.
2. La traduction se poursuit alors au-delà du cadre de lecture normal jusqu’à l’extrémité 3′ de l’ARNm et souvent jusque dans la queue poly(A), dont la succession d’adénosines est traduite en codons AAA codant pour la lysine, conduisant à la synthèse d’une queue polylysine aberrante à l’extrémité C-terminale de la protéine naissante.
Le ribosome finit alors par atteindre l’extrémité 3′ de l’ARN messager où il reste bloqué (ribosome stalling) avec l’ARNm, l’ARNt, et la chaîne polypeptidique encore attachée, en l’absence de signal normal de terminaison et de dissociation ribosomique ( terminaison de la traduction).
En association avec HBS1L, une GTPase homologue de eRF3, Pelota reconnaît les ribosomes bloqués et favorise le recrutement des mécanismes responsables de leur dissociation.
2. Ces facteurs favorisent notamment le recrutement de l'ATPase ABCE1, qui utilise l'énergie de l'hydrolyse de l'ATP pour dissocier le ribosome bloqué en sous-unités 40S et 60S distinctes (ribosome splitting), permettant leur recyclage (ribosome recycling) et le démantèlement du complexe traductionnel aberrant.
Schéma des voies NSD et NGD et du contrôle qualité associé aux ribosomes (RQC)
(Figure : vetopsy.fr d'après Ha et coll)
Ribosome Quality Control (RQC)
La grande sous-unité 60S restant associée à une chaîne polypeptidique incomplète est prise en charge par le complexe de contrôle qualité ribosomique (RQC).
a. LTN1 ubiquitine les régions accessibles de la chaîne polypeptidique incomplète encore associée à la sous-unité 60S, marquant ainsi cette protéine aberrante pour sa dégradation.
b. Lorsque l'ubiquitination est insuffisante ou retardée, NEMF peut également induire l'ajout d'extensions C-terminales riches en alanine et en thréonine appelées CAT tails (C-terminal alanine-threonine tails).
Ces extensions résultent d'un mécanisme particulier dans lequel NEMF recrute des ARNt chargés en alanine et en thréonine vers la sous-unité 60S encore associée à la chaîne polypeptidique incomplète.
Le centre peptidyl-transférase ajoute alors successivement ces résidus à l'extrémité C-terminale de la protéine sans lecture supplémentaire de l'ARN messager.
Les CAT tails facilitent ainsi l'accessibilité du substrat aux mécanismes d'ubiquitination et constituent un signal supplémentaire favorisant l'élimination des protéines aberrantes qui échappent au contrôle qualité initial.
2. Les protéines ubiquitinées sont ensuite prises en charge par VCP/p97, une ATPase de la famille AAA+, qui utilise l'énergie de l'hydrolyse de l'ATP pour extraire mécaniquement la chaîne polypeptidique de la sous-unité 60S avant son transfert vers le protéasome où elle sera dégradée.
Dégradation de l'ARN messager aberrant
En parallèle de la prise en charge de la protéine incomplète par le système RQC, l'ARN messager dépourvu de codon stop est dirigé vers les mécanismes cellulaires de dégradation des ARN.
1. La voie NGD (no-go decay) intervient lorsque la progression des ribosomes est fortement ralentie ou bloquée sur un ARN messager au cours de la traduction.
a. Contrairement à la voie NSD, les ARN ciblés par le NGD possèdent généralement un codon stop fonctionnel, mais la traduction ne peut pas se poursuivre correctement en raison d’un obstacle rencontré par les ribosomes, appelé blocage ribosomique (ribosome stalling).
b. Ces blocages peuvent être provoqués notamment par :
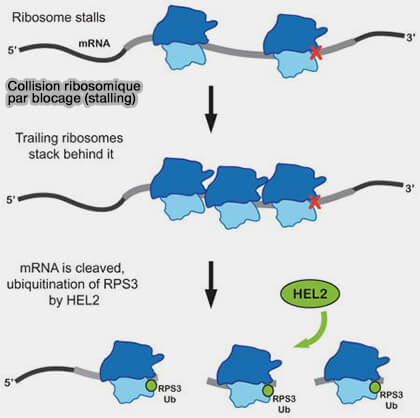
Blocage et collision ribosomique
(Figure : vetopsy.fr d'après Simms et coll)
des structures secondaires très stables des ARN,
des lésions chimiques ou des ARN endommagés,
des séquences difficiles à traduire, notamment certaines répétitions ou régions riches en codons rares,
des défauts de décodage ou certaines anomalies de maturation des ARN messagers.
a. Dans des conditions normales, plusieurs ribosomes peuvent traduire simultanément un même ARN messager sous forme de polysomes (ou polyribosomes), les ribosomes progressant à une vitesse relativement compatible et gardant une certaine distance entre eux le long de l’ARNm au cours de la synthèse protéique.
b. Lorsqu’un ribosome est fortement ralenti ou bloqué (leading/stalled ribosome), les ribosomes situés en amont continuent cependant d’avancer et peuvent finir par entrer en collision avec lui (trailing ribosomes), créant une interface structurale particulière entre deux ribosomes 80S.
La simple présence d’un ribosome bloqué n’est généralement pas suffisante pour déclencher la voie NGD.
Le signal principal correspond à l’accumulation de plusieurs ribosomes sur le même ARN messager et à leur collision, phénomène révélant un défaut persistant de traduction.
3. La collision ribosomique provoque une déformation locale des surfaces du ribosome, une exposition inhabituelle de certaines protéines ribosomiques et la formation d’une plateforme structurale absente lors d’une traduction normale.
a. Hel2 (ZNF598 chez les mammifères) est une E3 ubiquitine ligase qui ubiquitine certaines protéines ribosomiques de la 40S, situées à l’interface des ribosomes empilés, notamment uS10/RPS20, ainsi que dans certains contextes uS3/RPS3 et eS10/RPS10, ce qui transforme la collision en signal moléculaire détectable par les systèmes NGD/RQC.
b. Chez les mammifères, l'ubiquitination des protéines ribosomiques par ZNF598 favorise également le recrutement du complexe ASCC (Activating Signal Cointegrator Complex), notamment de l'hélicase ASCC3.
Ce complexe participe à la résolution des collisions ribosomiques et au démantèlement des complexes de traduction aberrants en coopération avec les mécanismes de sauvetage ribosomique et de contrôle qualité associés au NGD.
b. Le NGD déclenche alors le recrutement de facteurs spécialisés capables d’induire le clivage endonucléolytique de l’ARNm à proximité des ribosomes bloqués ( mécanismes de la voie NSD).
La dégradation des fragments d’ARN et le recyclage des ribosomes font ensuite intervenir plusieurs facteurs communs aux voies NGD et NSD.
Surveillance des autres ARN
La surveillance des autres ARN cellulaires concerne principalement les ARN impliqués dans les complexes ribonucléoprotéiques nucléaires, notamment :
1. Contrairement à la surveillance des ARN messagers, majoritairement couplée à la traduction cytoplasmique, ces mécanismes de contrôle qualité sont essentiellement associés aux étapes :
2. Les ARN présentant des défauts de maturation, de repliement, de modification chimique ou d’assemblage avec leurs protéines partenaires sont généralement :
retenus dans le noyau,
dirigés vers des mécanismes spécialisés de dégradation impliquant notamment le complexe de l'exosome nucléaire et différents complexes adaptateurs assurant la reconnaissance et l’adressage des ARN aberrants.




 codons et anticodons).
codons et anticodons).