L’ADN répétitif regroupe des séquences présentes en multiples copies dans le génome, organisées en répétitions en tandem ou en éléments dispersés, jouant des rôles structuraux, régulateurs et évolutifs.
Une part importante du génome est constituée de séquences répétées, regroupées sous le terme d’ADN répétitif.
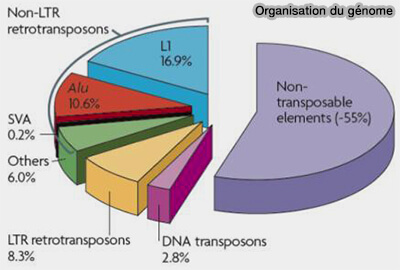
Organisation du génome (pourcentage)
(Figure : vetopsy.fr d'après Cordaux et Batzer)
Contrairement aux gènes codants, ces séquences ne produisent généralement pas de protéines, mais elles participent à l’organisation structurale des chromosomes, à la régulation de la chromatine et à l’évolution du génome.
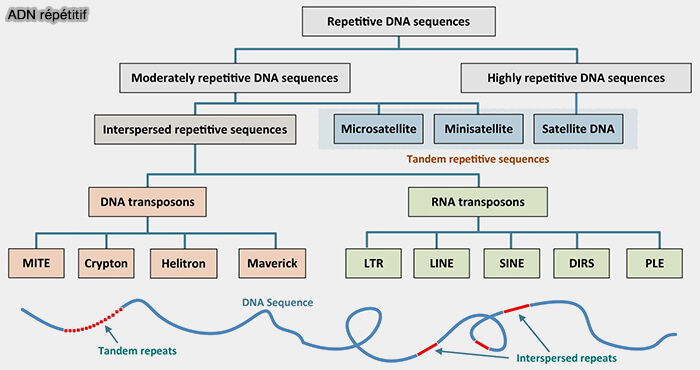
Selon leur organisation dans l’ADN, les séquences répétées sont classées en deux grandes catégories :
les répétitions en tandem, dans lesquelles les unités répétées sont disposées les unes à la suite des autres ;
les répétitions dispersées, correspondant principalement aux éléments transposables, capables de se multiplier et de se déplacer dans le génome.
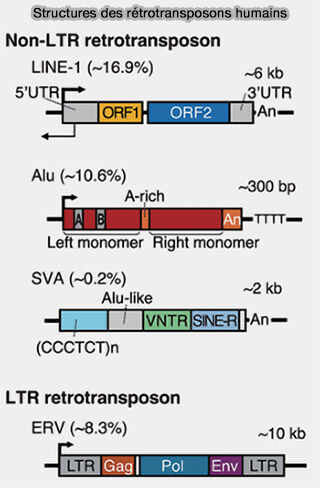
ADN répétitif
(Figure : vetopsy.fr d'après Liao et coll)
Répétitions en tandem
Les répétitions en tandem sont des séquences d’ADN répétées de manière adjacente, organisées en séries continues au sein du génome.
Selon la taille des unités répétées et l’organisation des répétitions, ces séquences sont classées en satellites, minisatellites et microsatellites.
En biologie moléculaire, une répétition en tandem signifie que les unités répétées sont disposées les unes à la suite des autres sur la molécule d’ADN, et non pas groupées par paires.
Gènes d'ADNr
Les gènes d’ADNr sont organisés en copies multiples en tandem, permettant une production élevée d’ARN ribosomiques (ARNr) nécessaire à la formation des ribosomes.
Les télomères sont constitués de séquences répétitives situées à l’extrémité des chromosomes eucaryotes.
Chez l’humain, ils sont constitués de répétitions en tandem de la séquence TTAGGG, formant une structure spécialisée qui protège les extrémités chromosomiques et empêche leur reconnaissance comme des cassures de l’ADN.
Les télomères sont étudiés dans une page spécifique.
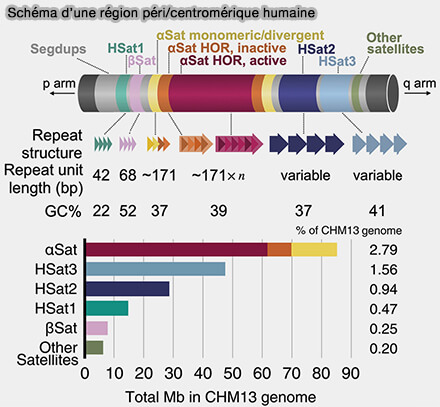
Schéma d'une région péri/centromérique humaine
(Figure : vetopsy.fr d'après Altemose et coll)
Anciennement, ces séquences étaient classées en satellites I, II et III, selon une classification fondée principalement sur leur densité en centrifugation CsCl et leur composition en bases (A classical revival: human satellite DNAs enter the genomics era 2023).
Aujourd’hui, les analyses génomiques permettent plutôt de distinguer deux grands ensembles fonctionnels.
2. Les séquences satellites péricentromériques comprennent plusieurs catégories.
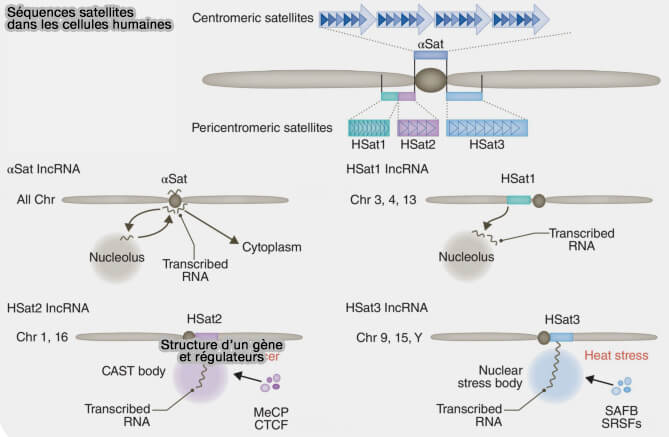
a. Les ADN satellites de type I (HSat1) sont constitués de répétitions en tandem relativement longues et hétérogènes, présentant une faible conservation de séquence et une organisation moins régulière.
Les ADN satellites de type I, principalement localisés dans les régions péricentromériques, participent surtout à l’organisation de l’hétérochromatine en recrutant des protéines comme HP1 et des complexes Polycomb, contribuant à une compaction stable, à la cohésion chromosomique et au maintien de l’intégrité structurale des régions centromériques adjacentes.
Séquences satellites dans les cellules humaines
(Figure : vetopsy.fr d'après Ninomiya et coll)
b. Les ADN satellites de type II (HSat2) correspondent à des répétitions en tandem plus courtes et plus homogènes, souvent riches en AT, organisées de manière plus régulière et compacte.
Ils favorisent la formation d’une hétérochromatine dense dans les régions péricentromériques.
c. Les ADN satellites de type III (HSat3) sont constitués de répétitions en tandem très homogènes, souvent formées de motifs courts répétés, organisées en structures hautement compactées au sein de l’hétérochromatine constitutive, notamment dans certaines régions péricentromériques spécifiques.
Ces séquences sont normalement silencieuses, mais peuvent être transcrites en ARN lors de conditions de stress cellulaire, contribuant à la formation des nuclear stress bodies.
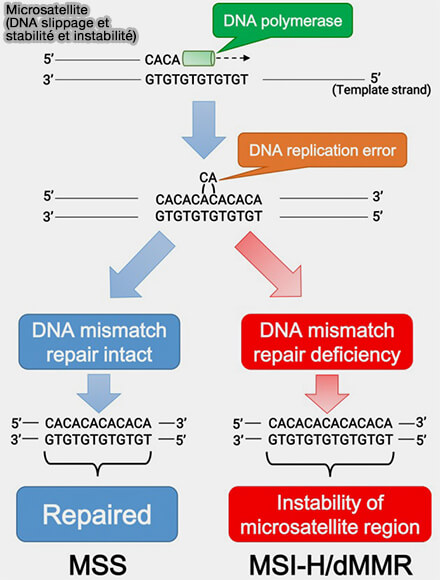
Microsatellites
Les microsatellites sont des courtes séquences répétées en tandem, généralement composées de motifs de 1 à 6 nucléotides, répétés plusieurs fois de suite, comme un motif CA, i.e. CACACACACA.
1. Leur variabilité provient principalement d’un mécanisme appelé glissement de la polymérase lors de la réplication de l’ADN (replication slippage).
Microsatellite (slippage)
(Figure : vetopsy.fr d'après Eso et coll)
Pendant la réplication, l’ADN polymérase copie une séquence déjà très répétitive, ce qui favorise des erreurs d’alignement entre le brin matrice et le brin en cours de synthèse.
Comme les motifs sont identiques ou presque identiques, le nouveau brin peut se décaler d’une ou plusieurs unités répétées sans que l’erreur soit immédiatement détectée.
2. Il peut alors se former des boucles transitoires :
sur le brin néosynthétisé, entraînant l’ajout d’une ou plusieurs unités répétées (expansion),
sur le brin matrice, entraînant la perte d’unités répétées (contraction).
Cette instabilité explique leur polymorphisme élevé, faisant des microsatellites des marqueurs génétiques utilisés pour l’identification, l’étude des populations et certaines analyses médicales, notamment :
en pathologie pour la détection de l’instabilité des microsatellites dans certains cancers, notamment les cancers colorectaux associés à un défaut du système de réparation des mésappariements.
Répétitions dispersées (éléments transposables)
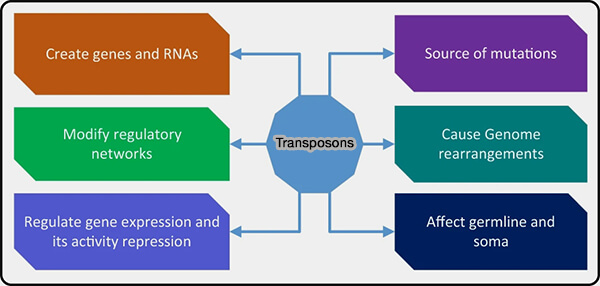
Les éléments transposables sont des séquences capables de se déplacer dans le génome.
Leur mobilité peut provoquer des insertions dans de nouveaux loci génomiques, susceptibles de perturber l’expression des gènes ou d’induire des mutations.
Pour limiter cette instabilité potentielle, les cellules ont développé plusieurs niveaux de contrôle, notamment :
une surveillance et une dégradation des ARN dérivés de transposons par des mécanismes de contrôle qualité des ARN, notamment la dégradation des ARN aberrants par le complexe de l’exosome nucléaire, les exoribonucléases cytoplasmiques de type XRN et certaines voies de surveillance telles que le nonsense-mediated decay ou NMD ( dégradation et surveillance des ARN).
1. On distingue deux grandes classes selon leur mécanisme de transposition.
les transposons à ADN,
les rétrotransposons.
Transposons
(Figure : vetopsy.fr d'après Liao et coll)
Chez l’homme, la majorité des éléments transposables actifs appartiennent aux rétrotransposons qui représentent environ 45 % du génome.
Rôles des transposons
(Figure : vetopsy.fr d'après Liao et coll)
Transposons à ADN
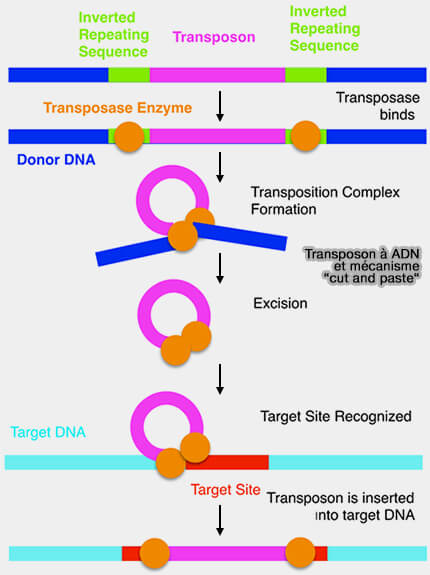
1. Les transposons à ADN se déplacent directement sous forme d’ADN par un mécanisme de type " couper-coller " catalysé par une transposase.
Transposons à ADN et mécanisme " cut and paste "
(Figure : vetopsy.fr d'après Alana Gyemi)
a. La transposase reconnaît des séquences répétées inversées terminales (Terminal Inverted Repeats, TIR) situées aux extrémités du transposon et rapproche ses deux extrémités au sein d'un complexe synaptique.
Elle excise ensuite l'élément du site d'origine en clivant les deux brins d'ADN, puis catalyse son insertion dans une nouvelle région du génome.
L'intégration s'effectue généralement au niveau d'une courte séquence cible. La transposase réalise des coupures décalées dans l'ADN cible, puis l'insertion du transposon est suivie par la réparation des brèches par les enzymes cellulaires, ce qui génère de part et d'autre du transposon des répétitions directes caractéristiques du site cible.
b. Contrairement aux rétrotransposons, ce mécanisme implique le déplacement direct de l’ADN sans intermédiaire ARN et ne conduit généralement pas à une augmentation du nombre de copies dans le génome.
Remarque : bien que la transposition provoque des déplacements et des réarrangements de séquences d'ADN, elle est généralement distinguée des mécanismes de recombinaison proprement dits, qui reposent sur l'échange ou le réarrangement contrôlé d'informations génétiques entre molécules d'ADN ( recombinaison).
2. Les transposons à ADN comprennent plusieurs familles (MITE, Helitron, Crypton, Maverick), définies par leur mécanisme de transposition et leur organisation génétique, mais la plupart sont aujourd’hui inactifs dans le génome humain.
Les rétrotransposons se déplacent par un mécanisme " copier-coller " impliquant un intermédiaire ARN.
Ce mécanisme, appelé Target-Primed Reverse Transcription (TPRT), permet l’intégration d’une nouvelle copie du rétrotransposon dans le génome sans excision de la copie initiale.
1. Les rétrotransposons sont d’abord transcrits en ARN de transposon qui joue un rôle analogue à celui d’un ARN messager (ARNm).
Il peut être traduit en protéines de transposon nécessaires à la rétrotransposition, notamment une endonucléase et une transcriptase inverse.
Les protéines produites s’associent à l’ARN de transposon pour former un complexe ribonucléoprotéique qui cible une nouvelle région du génome.
2. L’endonucléase clive alors l’ADN cible et génère une extrémité 3′ hydroxyle libre, qui sert d’amorce pour la synthèse d’ADN.
3. La transcriptase inverse utilise ensuite l’ARN de transposon comme matrice pour synthétiser une nouvelle copie d’ADN directement au site d’insertion.
Classification des rétrotransposons
Les rétrotransposons comprennent plusieurs classes définies par leur organisation structurale et leur mécanisme d’intégration notamment :
en rétrotransposons à LTR (Long Terminal Repeat), proches des rétrovirus endogènes, ainsi que d’autres groupes plus anciens et moins répandus chez les vertébrés, comme les DIRS et les PLE (Penelope-like elements).
en rétrotransposons non-LTR, principalement représentés par les LINE (Long Interspersed Nuclear Elements) et les SINE (Short Interspersed Nuclear Elements), qui constituent les formes dominantes d’éléments mobiles du génome chez l'homme
1. Les LINE, notamment LINE-1 chez l’humain, codent des protéines (endonucléase et reverse transcriptase) qui permettent leur insertion dans de nouvelles régions du génome, contribuant à la plasticité génomique mais pouvant également provoquer des mutations par insertion.
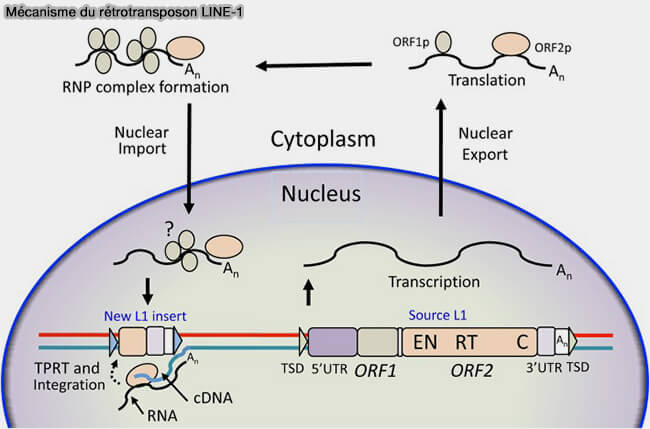
Mécanisme du rétrotransposon LINE-1
(Figure : vetopsy.fr d'après Lou et coll)
La structure d'un L1 complet comprend un 5′UTR, région non traduite contenant le promoteur du L1, ORF1, gène codant la protéine ORF1p, une protéine liant l’ARN, ORF2, gène codant la protéine ORF2p, qui possède deux activités enzymatiques EN (endonucléase) et RT (transcriptase inverse), 3′UTR, région non traduite, queue poly(A), TSD (Target Site Duplication), petites duplications de l’ADN cible générées lors de l’insertion.
2. Le mécanisme, appelé Target-Primed Reverse Transcription (TPRT), permet l’intégration d’une nouvelle copie du rétrotransposon dans le génome sans excision de la copie initiale.
Un élément LINE-1 actif est d’abord transcrit en ARN à partir de son promoteur, puis cet ARN est traduit dans le cytoplasme en deux protéines, ORF1p et ORF2p.
ORF1p est une protéine liant l’ARN, tandis que ORF2p possède une activité endonucléase et une activité transcriptase inverse.
L’endonucléase coupe alors l’ADN chromosomique et génère une extrémité 3′-OH libre servant d’amorce.
La transcriptase inverse utilise ensuite l’ARN LINE-1 comme matrice pour synthétiser une copie d’ADN directement au site d’insertion.
Remarque : la transcriptase inverse impliquée dans la rétrotransposition des LINE est la protéine ORF2p, alors que dans les télomères, la transcriptase inverse correspond à TERT, sous-unité catalytique de la télomérase.
SINE (Short Interspersed Nuclear Elements)
Les SINE (Short Interspersed Nuclear Elements), contrairement aux LINE, sont des éléments non autonomes, incapables de se mobiliser seuls, car ils ne codent pas ces enzymes et utilisent la machinerie enzymatique des LINE, en particulier LINE-1, pour leur rétrotransposition.
L'ARN 7SL est un ARN non codant constituant de la particule de reconnaissance du signal (SRP), un complexe ribonucléoprotéique qui intervient au cours de la traduction en dirigeant les protéines destinées à être sécrétées ou insérées dans les membranes vers le réticulum endoplasmique (RE).
Au cours de l'évolution, certaines copies ancestrales de ces ARN ont acquis la capacité de se rétrotransposer grâce aux protéines codées par les LINE, donnant naissance aux différentes familles de SINE.
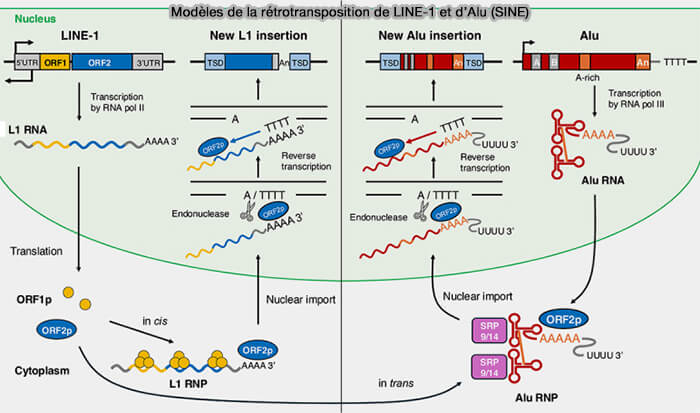
Modèles de la rétrotransposition de LINE-1 et d’Alu (SINE)
(Figure : vetopsy.fr d'après Luqman-Fatah et coll)
2. Chez l’humain, la famille la plus abondante est constituée des éléments Alu (~300 pb), dérivés d'un ARN 7SL ancestral.
Les éléments Alu sont des éléments transposables présents à plus d'un million de copies et représentent environ 10 % du génome humain, et qui, au cours de l'évolution, se sont accumulés principalement dans les introns et les régions non traduites des gènes, où ils participent aujourd'hui à la structuration du génome et à la régulation de l’expression génique.
L’ARN Alu est reconnu par les protéines LINE-1, qui réalisent la coupure de l’ADN cible et la rétrotranscription de l’ARN Alu, permettant son insertion dans le génome par le même mécanisme TPRT.
a. Lorsque deux séquences Alu sont présentes en orientation inverse dans un même gène, par exemple dans deux introns ou dans des régions non traduites (UTR), elles sont transcrites en même temps que le reste du gène dans un même pré-ARNm.
b. En raison de leur forte homologie de séquence, ces deux éléments Alu complémentaires peuvent alors s'apparier spontanément, formant une longue région d’ARN double brin, impliquée dans plusieurs mécanismes cellulaires pouvant :






 dégradation et surveillance des ARN).
dégradation et surveillance des ARN).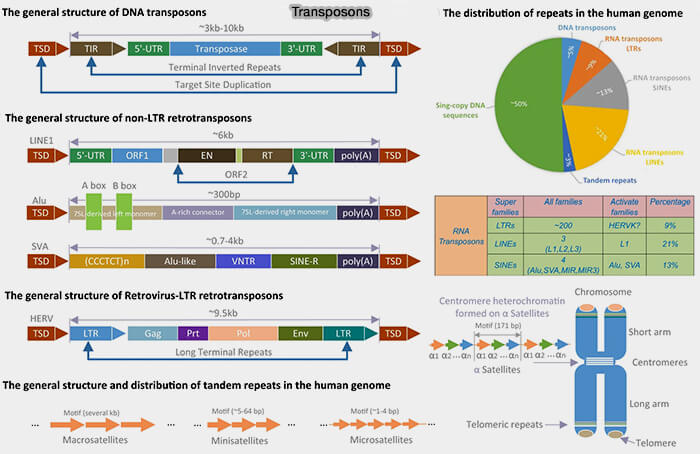
{kind=link}