


Acides nucléiques
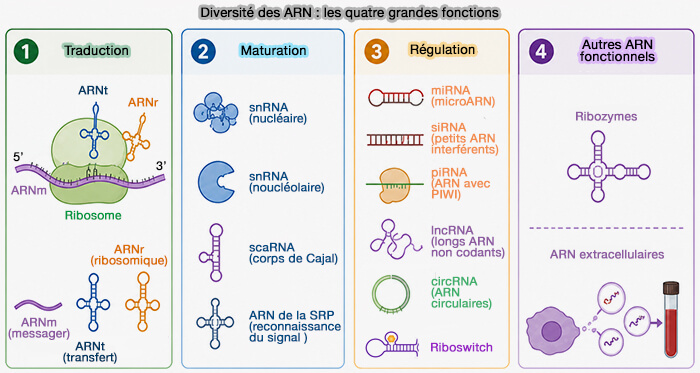
ARN : types et fonctions
ARN impliqués dans la maturation des ARN
snARN, snoARN et scaARN
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les snARN, snoARN et scaARN sont des ARN non codants spécialisés qui dirigent la maturation des ARN en assurant des modifications et des processus essentiels à l’expression génique.
Les ARN non codants impliqués dans la maturation des ARN constituent une catégorie spécialisée de petits ARN fonctionnels qui interviennent dans les différentes étapes de transformation des transcrits primaires en ARN matures.
(Figure : vetopsy.fr)
Plusieurs classes d’ARN non codants s’associent à des complexes ribonucléoprotéiques qui reconnaissent les ARN cibles et catalysent ou guident les réactions de maturation des ARN nucléaires (RNA processing) impliqués dans l’expression génique.
- Les snARN (small nuclear RNA) sont les composants essentiels du spliceosome impliqués dans l’’épissage des pré-ARNm.
- Les snoARN (small nucleolar RNA) sont des ARN guides qui dirigent les modifications chimiques des ARN ribosomiques (ARNr) dans le nucléole.
- Les scaARN (small Cajal body-specific RNA) sont ARN apparentés aux snoARN localisés dans les corps de Cajal, responsables des modifications post-transcriptionnelles des snARN spliceosomaux.
snARN (small nuclear RNA))
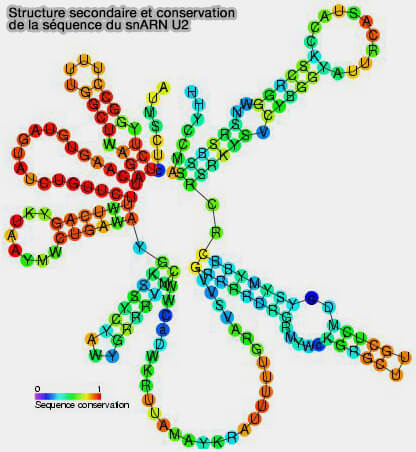
Les snARN (small nuclear RNA) sont une classe de petits ARN non codants localisés dans le noyau, dont la taille varie généralement entre 100 et 300 nucléotides.
Structure et classification
(Figure : vetopsy.fr d'après vieterre.fr)
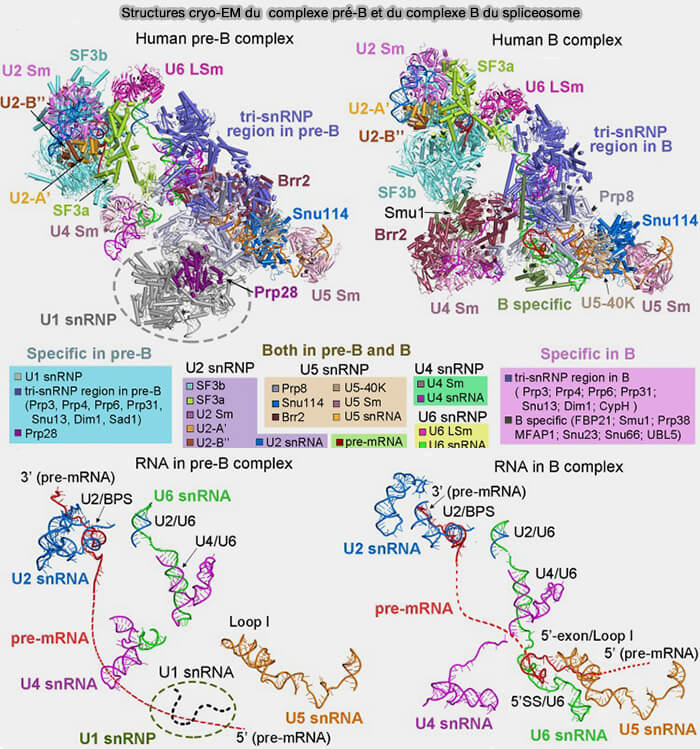
1. Les snARN adoptent généralement une structure secondaire organisée en tiges-boucles intramoléculaires et s’associent à des protéines Sm pour former le cœur des ribonucléoprotéines nucléaires appelées snRNP (small nuclear ribonucleoproteins), qui constituent les composants structuraux et fonctionnels du spliceosome.

Vous pouvez lire : Structures of the human pre-catalytic spliceosome and its precursor spliceosome (2018).
Ils sont désignés selon leur coefficient de sédimentation en unités de Svedberg (U1, U2, U4, U5, U6 et U7) et jouent un rôle essentiel dans la maturation des transcrits primaires.
2. Les snARN U1, U2, U4, U5 et U6 sont transcrits par l’ARN polymérase II et constituent les composants ARN essentiels du spliceosome.
Ils assurent des fonctions complémentaires.
- Le snARN U1 reconnaît le site d’épissage 5′.
- Le snARN U2 se fixe au point de branchement de l’intron.
- Les snARN U4 et U6 forment initialement un duplex qui régule l’activation catalytique du spliceosome.
Au cours de l’activation du spliceosome, U4 est libéré et le snARN U6 s’apparie avec U2 pour former le centre catalytique ARN de la machinerie d’épissage.
- Le snARN U6 participe directement à la catalyse de la réaction d’épissage.
- Le snARN U5 intervient dans l’alignement des exons, permettant leur ligature après excision de l’intron.
(Figure : vetopsy.fr d'après Zhan et coll)
Remarque : le snARN U7, distinct des snARN spliceosomaux, intervient dans la maturation des ARN d’histones, un processus spécifique des Histone Locus Bodies ( mécanisme complet de maturation de ces transcrits).
mécanisme complet de maturation de ces transcrits).
Il reconnaît une séquence spécifique située en aval de la région codante des pré-ARNm d’histones et recrute les facteurs nécessaires au clivage de l’extrémité 3′, générant les ARNm d’histones matures dépourvus de queue poly(A).
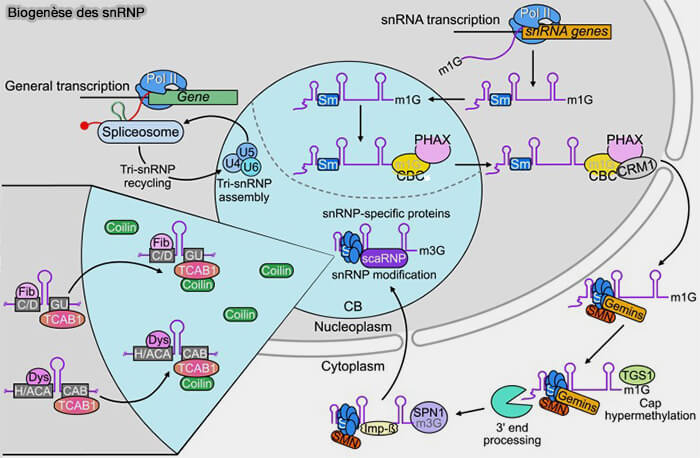
Maturation des snARN
Les snARN subissent plusieurs étapes de maturation post-transcriptionnelle conduisant à la formation des snRNP fonctionnels impliqués dans l’épissage des pré-ARNm.
1. Dans le noyau, les snARN synthétisés par l’ARN polymérase II :
- acquièrent d’abord une coiffe 5′ monométhylguanosine (m7G), reconnue par le complexe CBC (Cap-Binding Complex),
- puis subissent un clivage de leur extrémité 3′ par le complexe Integrator, associé au domaine C-terminal (CTD) de l’ARN polymérase II, étape essentielle à leur stabilité et à leur biogenèse.
(Figure : vetopsy.fr d'après Taliansky et coll)
2. Les pré-snARN sont ensuite exportés transitoirement vers le cytoplasme, notamment grâce à l’adaptateur d’export PHAX (Phosphorylated Adapter RNA Export Protein) et à l’exportine CRM1/XPO1 ( export nucléocytoplasmique).
- Les snARN s’assemblent avec les protéines Sm pour former le cœur ribonucléoprotéique des futurs snRNP.
- Cet assemblage favorise notamment la conversion de la coiffe m7G en coiffe triméthylguanosine (TMG), caractéristique de nombreux snRNP du spliceosome.
3. Les snRNP nouvellement assemblés sont ensuite réimportés dans le noyau via la snurportine et l’importine-β ( import nucléocytoplasmique).
4. Les snRNP subissent différentes étapes de maturation finale, notamment la 2′-O-méthylation et la pseudouridylation guidées par les scaARN dans les corps de Cajal compartiments nucléaires spécialisés dans la maturation des snRNP avant leur participation à l’épissage.
snoARN (small nucleolar RNA)
Les snoARN (small nucleolar RNA) sont de petites molécules d’ARN non codants localisées principalement dans le nucléole, où se déroule une grande partie de la maturation des ARN ribosomiques (ARNr).
Leur taille varie généralement entre 60 et 300 nucléotides et ils s’associent à des protéines spécifiques pour former des complexes ribonucléoprotéiques appelés snoRNP (small nucleolar ribonucleoprotein).
Structure et classification
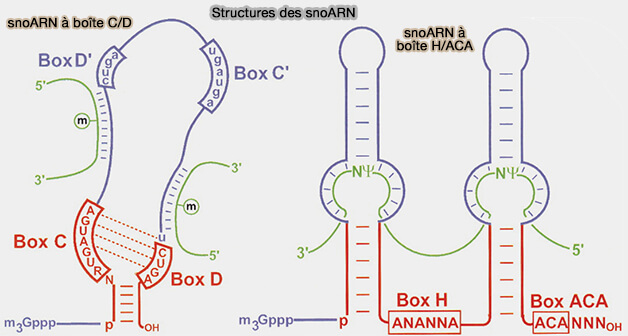
Les snoARN sont classés en deux grandes familles structurales selon leurs motifs de séquence conservés et le type de modification qu’ils dirigent (Small Nucleolar RNAs An Abundant Group of Noncoding RNAs with Diverse Cellular Functions 2002).
1. Les snoARN de type C/D contiennent deux motifs conservés appelés boîte C (RUGAUGA) et boîte D (CUGA), et possèdent souvent une seconde paire interne de motifs appelée boîtes C′ et D′, structurées de manière analogue aux boîtes C et D et permettant la formation d’un second site guide au sein du snoARN (Emerging Data on the Diversity of Molecular Mechanisms Involving C/D snoRNAs 2021).
a. Ces deux motifs sont rapprochés dans la structure tridimensionnelle de l’ARN et participent à la formation d’une organisation en tige-boucle qui permet :
- le recrutement des protéines spécifiques du complexe snoRNP,
- le rapprochement des extrémités 5′ et 3′ du snoARN et la stabilisation de la structure du complexe.
b. Les snoARN de type C/D guident la méthylation en position 2’-O du ribose de nucléotides spécifiques des ARN cibles, réaction enzymatique étant catalysée par la fibrillarine associée au complexe snoRNP, qui comprend également les protéines NOP56, NOP58 et SNU13 assurant la stabilité et l’organisation du complexe.
(Figure : vetopsy.fr d'après Kiss)
2. Les snoARN de type H/ACA possèdent une organisation structurale différente des snoARN de type C/D (Emerging Data on the Diversity of Molecular Mechanisms Involving C/D snoRNAs 2021)
a. On note la présence d’une boîte H interne, de séquence consensus ANANNA, et d’une boîte ACA située près de l’extrémité 3′ de l’ARN.
Les ARN adoptent généralement une structure constituée de deux tiges-boucles séparées par une région charnière contenant la boîte H, tandis que la boîte ACA se situe à quelques nucléotides de l’extrémité 3′.
b. Les snoARN de type H/ACA guident la pseudouridylation dans les ARN cibles, réaction catalysée par la dyskérine associée au complexe snoRNP, qui comprend également les protéines NOP10, NHP2 et GAR1 participant à la stabilité et au positionnement du complexe.
3. Les snoARN possèdent des régions appelées séquences guides, constituées de segments d’ARN capables de s’apparier par complémentarité de bases avec une région spécifique de l’ARN cible pour positionner précisément le nucléotide à modifier en face du site catalytique du complexe snoRNP (SnoRNAs: Exploring Their Implication in Human Diseases 2024).
- Dans les snoARN de type C/D, la base de l’ARN cible qui sera méthylée est généralement située cinq nucléotides en amont de la boîte D du snoARN, ou de la boîte D′ lorsqu’un second site guide est présent.
- Dans les snoARN de type H/ACA l’uridine à modifier est placée dans une poche structurale formée par les deux tiges-boucles du snoARN.
Biogenèse et maturation des snoARN
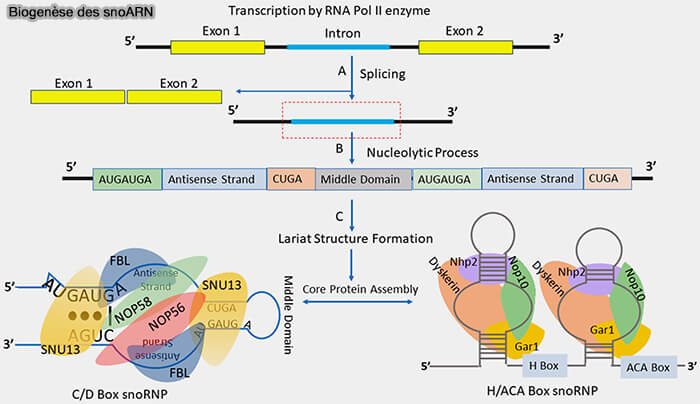
1. Chez les vertébrés, la majorité des snoARN ne sont pas transcrits à partir de gènes indépendants.
a. Leurs séquences sont généralement localisées dans les introns de gènes appelés gènes hôtes (host genes), souvent impliqués dans la biogenèse des ribosomes, la traduction ou d’autres fonctions nucléaires (SnoRNAs: Exploring Their Implication in Human Diseases 2024).
- Ces gènes sont transcrits par l’ARN polymérase II pour produire un pré-ARNm contenant les séquences des futurs snoARN.
- Lors de l’épissage du pré-ARNm, les introns contenant les séquences des snoARN sont excisés.
- Au lieu d’être entièrement dégradés, certains de ces introns sont pris en charge par des mécanismes de maturation spécifiques qui permettent la libération et la stabilisation des snoARN qu’ils contiennent.
b. Ainsi, le précurseur immédiat de nombreux snoARN n’est pas un transcrit indépendant mais un intron issu du pré-ARNm d’un gène hôte.
(Figure : vetopsy.fr d'après Chauhan et coll)
2. Après leur libération à partir des introns des gènes hôtes ou de précurseurs indépendants,leurs extrémités 5′ et 3′ subissent différentes étapes de maturation.
- La maturation de l’extrémité 3′ fait notamment intervenir plusieurs ribonucléases nucléaires, dont l’exosome nucléaire et ses sous-unités catalytiques EXOSC10/RRP6 et DIS3, conduisant à la formation de snoARN de longueur définie.
- Selon leur mode de biogenèse, certains snoARN peuvent acquérir à leur extrémité 5′ une coiffe triméthylguanosine (m3GpppN) caractéristique de nombreux petits ARN nucléaires et nucléolaires, qui correspond à une guanosine triphosphate hyperméthylée impliquée dans la stabilité de l’ARN et son trafic nucléaire.
3. Les snoARN s’assemblent ensuite progressivement avec des protéines spécifiques pour former des complexes snoRNP stables.
- Les snoARN de type C/D recrutent notamment la fibrillarine, NOP56, NOP58 et SNU13, tandis que les snoARN de type H/ACA s’associent à la dyskérine, NOP10, NHP2 et GAR1.
- L’assemblage de ces protéines favorise le repliement structural des snoARN, protège leurs extrémités contre la dégradation et permet la formation de complexes snoRNP fonctionnels
4. Les snoRNP matures s’accumulent principalement dans le nucléole où elles participent à la maturation des ARN ribosomiques, notamment par le guidage des 2′-O-méthylations et des pseudouridylations.
scaARN (small Cajal body-specific RNA)
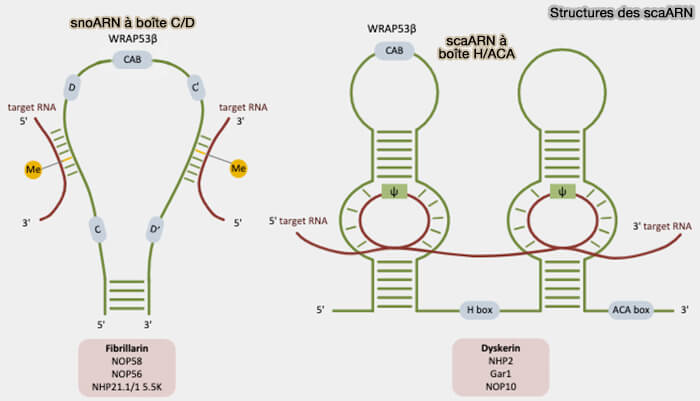
Les scaARN (small Cajal body-specific RNA) sont une classe spécialisée de petits ARN non codants étroitement apparentés aux snoARN (small nucleolar RNA).
Alors que les snoARN interviennent principalement dans la maturation des ARN ribosomiques au sein du nucléole, les scaARN sont localisés dans les corps de Cajal où ils participent principalement à la maturation des snARN du spliceosome.
1. Comme les snoARN, les scaARN :
- appartiennent aux deux grandes classes structurales C/D et H/ACA.
- subissent des mécanismes de biogenèse et de maturation similaires conduisant à la formation de complexes scaRNP fonctionnels (biogenèse et maturation des snoARN),
- s’associent à des protéines spécifiques pour former des complexes ribonucléoprotéiques guides appelés scaRNP.
2. Les scaARN possèdent des séquences guides constituées de segments d’ARN capables de s’apparier par complémentarité de bases avec les snARN cibles, permettant de positionner précisément les nucléotides à modifier au niveau du site catalytique des complexes scaRNP.
- Ces séquences guides ne sont pas modifiées elles-mêmes, mais dirigent la maturation post-transcriptionnelle des snARN spliceosomaux, i.e. U1, U2, U4, U5 et U6 qui participent tous à l’épissage, notamment par la 2′-O-méthylation et la pseudouridylation.
- Ces modifications contribuent à la stabilité structurale des snARN et au fonctionnement correct du spliceosome lors de la reconnaissance des sites d’épissage et de la catalyse des réactions d’épissage ( épissage).

Les scaARN, leurs complexes ribonucléoprotéiques (scaRNP) et leur rôle sont étudiés dans le chapitre des corps de Cajal.
(Figure : vetopsy.fr adaptée d'après Coucoravas)
ARN non codants impliqués dans la régulation des ARN
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation