Le code des histones correspond à l’ensemble des modifications post-traductionnelles des histones et à leur combinaison, qui modulent l’organisation de la chromatine et l’expression des gènes.
Principe du code des histones
Les modifications post-traductionnelles des histones constituent un mécanisme central de régulation de la chromatine, en en modifiant les interactions entre l’ADN et les histones ainsi que les interactions entre nucléosomes, ce qui module localement l’organisation de la chromatine et l’accessibilité de l’ADN aux complexes chromatiniens.
Ces modifications modulent l’accessibilité de l’ADN aux protéines impliquées dans la transcription, la réplication, la réparation de l’ADN et - l’organisation et la dynamique de la chromatine..
Principe combinatoire des modifications d’histones
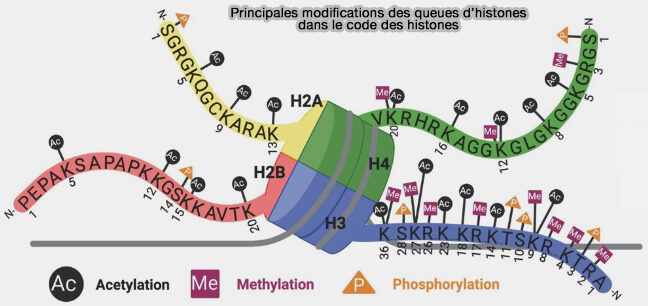
Principales modifications des queues des histones dans le code des histones
(Figure : vetopsy.fr d'après Malone et coll)
a. Les résidus les plus fréquemment modifiés sont notamment les lysines, arginines et sérines des histones H3 et H4, mais des modifications peuvent également toucher H2A et H2B.
b. Les modifications des histones comprennent plusieurs types de modifications covalentes : acétylation, méthylation, phosphorylation, ubiquitination, SUMOylation, ADP-ribosylation et citrullination ( cf. tableau).
Chaque modification peut modifier les propriétés physico-chimiques de la chromatine ou créer des sites de reconnaissance pour d’autres protéines.
2. Le code des histones repose sur un principe de combinaison de marques.
Une modification isolée possède souvent un effet limité, mais l’association de plusieurs modifications sur un même nucléosome ou sur des nucléosomes voisins constitue un signal moléculaire interprété par la cellule.
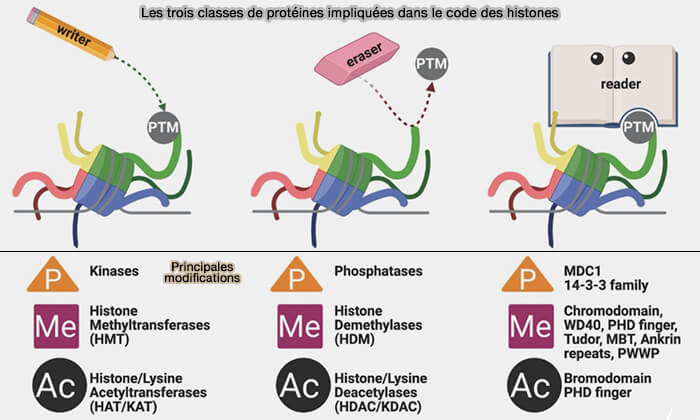
3. L’établissement et l’interprétation du code des histones impliquent trois grandes catégories de protéines qui permettent une régulation rapide et réversible de l’organisation de la chromatine :
des protéines writers, qui ajoutent les modifications,
des protéines erasers, qui les retirent,
des protéines readers, qui reconnaissent les marques et recrutent d’autres complexes chromatiniens.
Les trois classes de protéines impliquées dans le code des histones
(Figure : vetopsy.fr d'après Malone et coll)
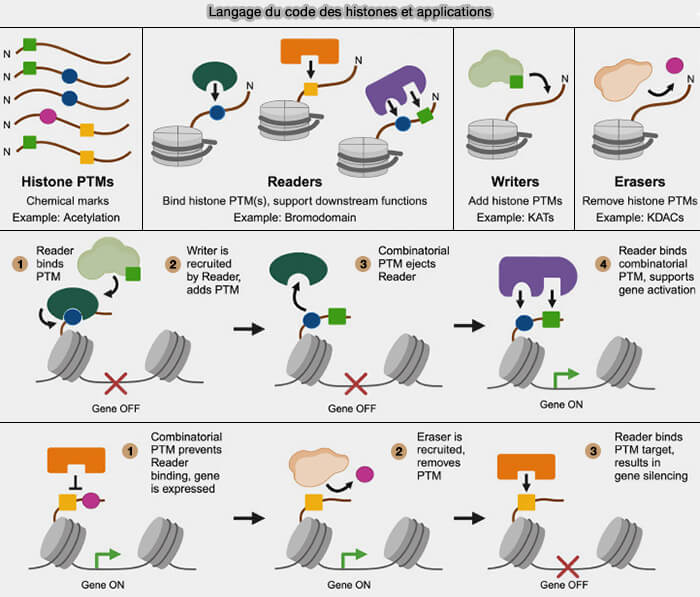
Interprétation du code des histones
Les modifications des histones ne modifient pas uniquement les propriétés structurales de la chromatine, mais constituent également des signaux moléculaires reconnus par des protéines lectrices.
Principe général de la lecture du code des histones
stabiliser des états chromatiniens actifs ou répressifs, contribuant à l’établissement et au maintien de domaines d’euchromatine ou d’hétérochromatine
3. Certaines protéines lectrices peuvent reconnaître simultanément plusieurs modifications des histones, ce qui permet d’intégrer différentes marques épigénétiques et de traduire leur combinaison en états fonctionnels distincts de la chromatine.
4. Le code des histones constitue ainsi un niveau majeur de régulation épigénétique capable de moduler l’expression des gènes sans modifier la séquence de l’ADN.
Langage du code des histones et applications
(Figure : vetopsy.fr d'après Weinzapfel et coll)
Domaines de reconnaissance des histones acétylées et méthylées
Les domaines de reconnaissance du code des histones ont été principalement caractérisés pour l’acétylation et la méthylation et sont détaillés ci-dessous.
Toutefois, d’autres modifications des histones peuvent également être reconnues par des protéines spécialisées, qui sont signalées dans les chapitres correspondants.
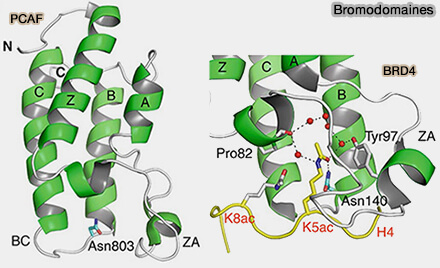
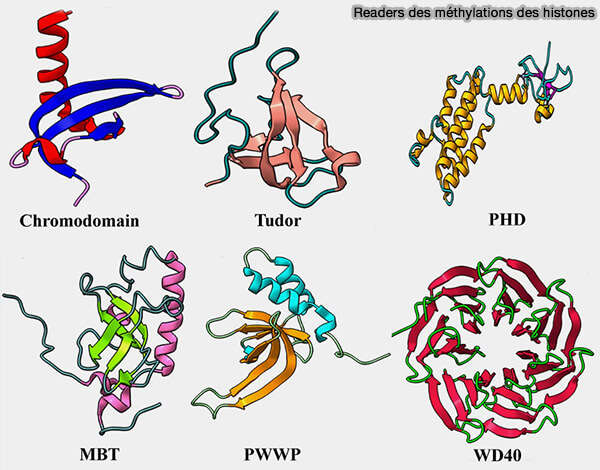
1. Les bromodomaines sont des modules structuraux d’environ 110 acides aminés formés par un faisceau de quatre hélices α qui délimitent une poche hydrophobe capable de reconnaître les lysines acétylées des histones (Bromodomain biology and drug discovery 2020).
Bromodomaines
(Figure : vetopsy.fr d'après Zaware et Zhou)
La reconnaissance repose sur l’interaction entre la lysine acétylée et cette poche structurale, stabilisée par des liaisons hydrogène et des interactions hydrophobes.
Les bromodomaines sont présents dans de nombreuses protéines régulatrices de la transcription, comme les protéines BET (BRD2, BRD3, BRD4), et participent au recrutement de complexes coactivateurs sur les régions de chromatine transcriptionnellement actives.
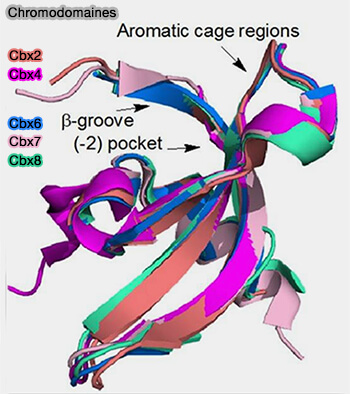
Ils possèdent généralement une cage aromatique formée de résidus aromatiques capables de reconnaître les lysines méthylées des histones par interactions cation-π.
Remarque : les protéines possédant un chromodomaine comprennent notamment les protéines HP1 reconnaissant principalement H3K9me2/3 associée à l’hétérochromatine ainsi que certaines protéines CBX du complexe Polycomb PRC1 reconnaissant principalement H3K27me3.
b. Les Tudor domains appartiennent à une large famille de domaines structuraux dérivés du motif Tudor, formés par un repliement en feuillet β antiparallèle qui crée une surface de reconnaissance pour les résidus méthylés.
Comme les chromodomaines, ils possèdent souvent une cage aromatique capable de reconnaître des lysines ou des arginines méthylées.
Ces domaines sont présents dans plusieurs protéines impliquées dans la régulation de la chromatine, la réparation de l’ADN ou la régulation transcriptionnelle.
c. Les PHD fingers (Plant Homeodomain fingers) sont de petits domaines riches en cystéines et histidines capables de coordonner deux ions zinc.
Ils adoptent une structure compacte stabilisée par ces ions métalliques et possèdent une surface de reconnaissance spécifique de certaines modifications histoniques.
Plusieurs PHD fingers reconnaissent spécifiquement la lysine triméthylée H3K4me3, marque caractéristique des promoteurs actifs, et participent au recrutement de complexes transcriptionnels.
d. Les domaines MBT (Malignant Brain Tumor domains) sont des modules structuraux d’environ 100 acides aminés appartenant à une famille de protéines associées à la chromatine.
Ces domaines possèdent une poche de liaison plus étroite capable de reconnaître préférentiellement les lysines mono- ou diméthylées des histones, plutôt que les triméthylées.
La reconnaissance repose sur une cavité structurale relativement étroite qui limite l’accès aux formes fortement méthylées, ce qui confère aux domaines MBT une certaine sélectivité pour les états de méthylation faibles.
Les protéines contenant des domaines MBT sont souvent impliquées dans la répression transcriptionnelle et l’organisation de la chromatine, notamment dans des complexes régulateurs associés à la différenciation cellulaire et au maintien de l’identité cellulaire.
Remarque : les domaines MBT sont aussi appelés MBT repeat, MBT repeat domain, MBT-containing proteins, L3MBTL proteins (famille humaine).
Readers des méthylations des histones
(Figure : vetopsy.fr d'après Sehrawat et coll)
e. Les domaines PWWP (Pro-Trp-Trp-Pro) sont des modules structuraux appartenant à la famille des domaines de type Royal.
Ils possèdent un repliement caractéristique composé d’un feuillet β associé à un faisceau d’hélices α, formant une surface de reconnaissance pour certaines lysines méthylées des histones.
Ils contiennent généralement une cage aromatique permettant la reconnaissance de résidus méthylés par interactions cation-π.
Les domaines PWWP reconnaissent préférentiellement les marques H3K36me2 et H3K36me3 et sont présents dans plusieurs protéines impliquées dans l’organisation de la chromatine, l’élongation transcriptionnelle et la réparation de l’ADN.
f. Les domaines WD40 sont des domaines structuraux constitués de répétitions WD formant une structure en hélice β-propeller, généralement composée de plusieurs lames de feuillets β disposées de manière radiale autour d’un axe central.
Cette architecture crée une large surface d’interaction capable de reconnaître des séquences peptidiques spécifiques des histones modifiées.
Dans plusieurs complexes épigénétiques ,notamment les complexes COMPASS/MLL et Polycomb (PRC2), ces domaines participent à la reconnaissance de certaines marques histoniques, notamment H3K4me3 ou H3K27me3, et facilitent l’assemblage de complexes multiprotéiques impliqués dans la régulation transcriptionnelle.
Les protéines contenant des domaines WD40 jouent ainsi un rôle important comme plateformes d’interaction protéine-protéine, permettant l’intégration des signaux épigénétiques et la coordination de différentes activités enzymatiques au niveau de la chromatine.
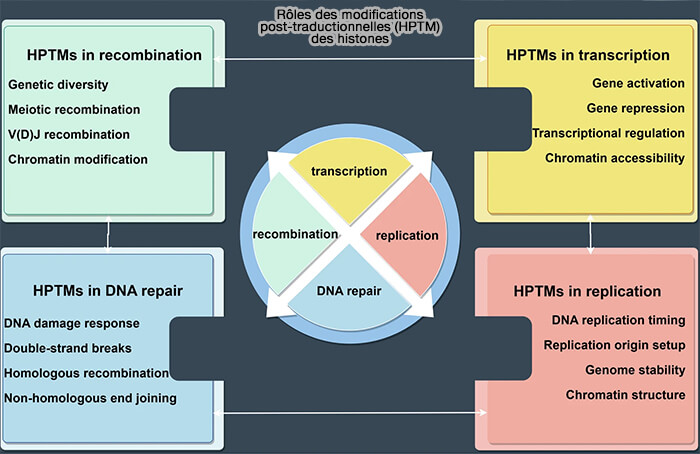
Rôles des modifications post-traductionnelles (HPTM) des histones
(Figure : vetopsy.fr d'après Yao et coll)
Principales modifications post-traductionnelles des histones
Les histones peuvent subir de nombreuses modifications post-traductionnelles qui modulent l’organisation de la chromatine et l’activité des gènes. Les principales modifications connues et leurs fonctions générales sont résumées dans le tableau suivant, chacune faisant l’objet d’un développement dans les pages correspondantes.




 cf. tableau).
cf. tableau).