


Lipides
Trafic non vésiculaire
LTP ou protéines de transfert des lipides
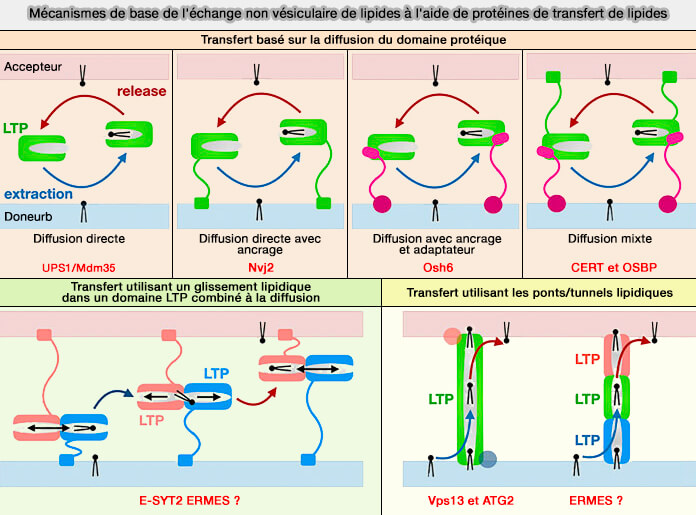
Mécanismes
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les mécanismes utilisés par les protéines de transfert des lipides (LTP ou Lipid Tranfer Protein) peuvent être grossièrement divisés en trois classes.
(Figure : vetopsy.fr d'après Chiapparino et coll)
Mécanisme général des LTP
Les LTP intracellulaires se localisent sur les sites de contact membranaire (MCS) au sein desquelles les LTP peuvent transférer les lipides d'un compartiment donneur à un compartiment accepteur (Lipid transfer proteins: the lipid commute by shuttles, bridges and tubes 2019)

Les chapitres sur les généralités des LTP sont largement inspirés de l'excellent article : Mechanisms of Non-Vesicular Exchange of Lipids at Membrane Contact Sites: Of Shuttles, Tunnels and, Funnels (2021).
1. Les mécanismes utilisés peuvent être grossièrement divisés en trois classes. Par contre, leurs frontières sont floues car certains systèmes d’échange lipidique en combinent plusieurs.
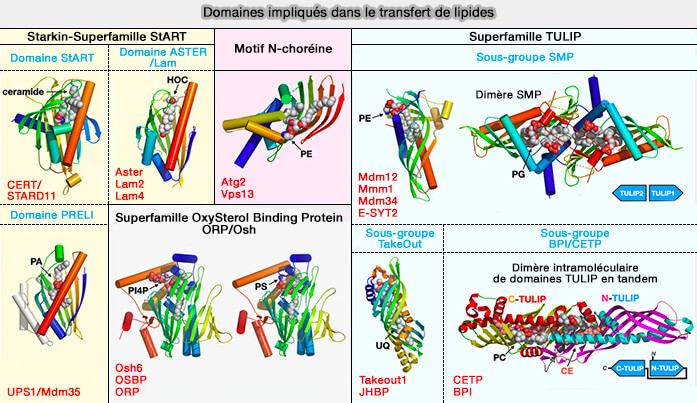
a. Le plus commun est le transfert lipidique par des navettes, i.e. transfert lipidique avec généralement un domaine de transfert lipidique d'environ 150 à 300 acides aminés.
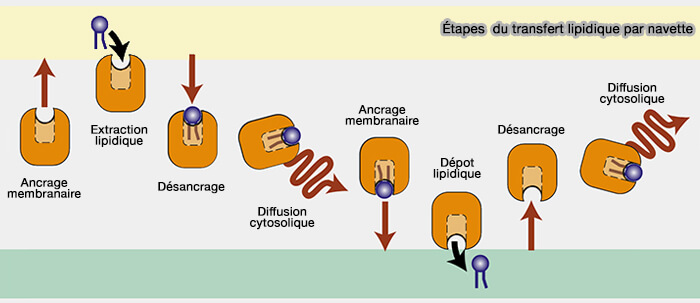
(Figure : vetopsy.fr d'après Wong et coll)
b. D'autres LTP sont de forme tubulaire avec un tunnel hydrophobe apparemment adapté à la canalisation des lipides d'une extrémité à l'autre par glissement lipidique.
- Les domaines SMP de la famille TULIP semble posséder un tunnel hydrophobe apparemment adapté à la canalisation des lipides d'une extrémité à l'autre par glissement lipidique.
- On peut aussi ranger dans cette catégorie le complexe Lpt des bactéries.
c. Les LTP en forme de ponts ou de tunnels sont des protéines beaucoup plus grandes comme Vps13 ou Atg2.
Remarque : cette classification peut être modifiée selon les auteurs, i.e. les TULIP ou le complexe LTP de la levure par exemple, peuvent être considérés comme des ponts entre les membranes.

Les deux dernières classes, mais surtout la dernière, ont l'énorme avantage de déplacer le lipide en l'absence partiel ou total de mouvement des protéines, ce qui permet le passage " en masse " (bulk en anglais) des lipides nécessaires à l’alimentation des structures membranaires dynamiques comme celles les mitochondries par exemple.
2. Un autre mécanisme particulier est le transfert de triglycérides (TG) par les seipines lors de la formation des goutelettes lipidiques (LD ou Lipid Droplet).
(Figure : vetopsy.fr d'après Egea)
3. En règle générale, ce qui est le cas du transfert lipidique par navette, transfert le plus fréquent, le lipide est inséré dans la cavité hydrophobe de la LTP avec une stoechiométrie 1:1 (Conformational dynamics of lipid transfer domains provide a general framework to decode their functional mechanism 2023).
- La sélection du lipide s'effectue par son groupe de tête et sa chaîne acyle (R-C(=O)-) grâce à une interaction spécifique avec des résidus distincts dans la cavité.
- Toutefois, certains LTP peuvent se lier à deux lipides à groupes de tête différents pour effectuer des échanges.
Remarque : dans le transfert par pont lipidique, la LTP peut héberger plus de 10 lipides.
(Figure : vetopsy.fr d'après Egea)
Quid du coût énergétique et de la direction du transfert ?
La plupart des LTP sont passives et ne dépensent pas directement d'énergie, i.e. hydrolyse des nucléotides ou gradient de potentiel membranaire.
1. Les enzymes responsables d'étapes spécifiques de la synthèse/transformation ou du catabolisme des lipides sont distribuées dans des organites spécifiques, i.e. les réactions métaboliques ayant lieu entre les compartiments peuvent entraîner un échange de lipides médié par la LTP le long de gradients de concentration (Phosphatidylserine flux into mitochondria unveiled by organelle-targeted Escherichia coli phosphatidylserine synthase PssA 2020).
- Par exemple, le transport des céramides par CERT (CERamide Transfer protein) du réticulum endoplasmique (RE) à l'appareil de Golgi est piloté par la transformation du céramide en sphingomyéline dans le Golgi qui ne peut pas être transporté en retour.
- La croissance des goutelettes lipidiques (LD ou Lipid Droplet) est le résultat des activités de la synthèse de triglycérides (TG) dans le RE et des enzymes attachées ou incorporées à la membrane des LD.
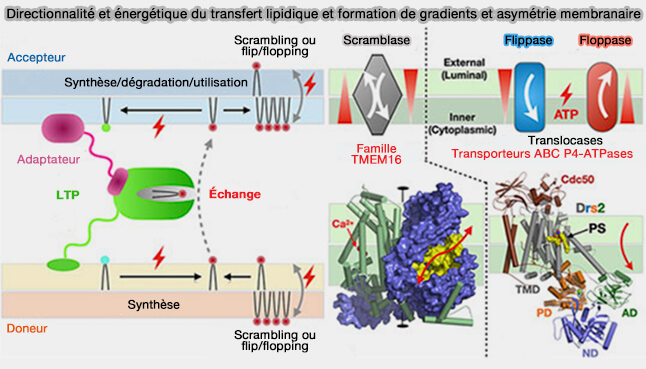
formation de gradients et asymétrie membranaire
(Figure : vetopsy.fr d'après Egea)
2. Deux classes de protéines membranaires intégrales participent au transfert des lipides d'un feuillet à un autre au sein d'une même membrane en modifiant l'asymétrie membranaire.
a. Les flippases/floppases, comme les transporteurs ABC ou les P4-ATPases, créent cette asymatrie en utilisant l'hydrolyse de l'ATP.
- Elles agissent au niveau des MCS et à l'intérieur de la cellule par la formation de gradients entre les feuillets au sein d'une membrane.
- Elles forment de nanodomaines latéraux non seulement au sein de la membrane, mais également au sein des feuillets (Transport Pathways That Contribute to the Cellular Distribution of Phosphatidylserine 2021).
b. Les scramblases abolissent l'asymétrie en équilibrant les distributions lipidiques entre les feuillets.
Elles sont indépendantes de l'ATP et agissent suivant des gradients de potentiel énergétique associés à un simple gradient de concentration chimique ou au potentiel électrique membranaire (résultant de distributions de charges).
3. Un autre mécanisme utilise l'ATP pour l'échange de lipides au niveau d'un site de contact membranaire en utilisant l'énergie du gradient chimique, comme pour OSBP ou Osh6.
- La biosynthèse de PI(4)P au niveau de l'appareil de Golgi est sous le contrôle de l'hydrolyse de l'ATP.
- L'OSBP a une affinité plus élevée pour le PI(4)P que pour le cholestérol, donc son échange contre le PI(4)P est facilité au niveau du Golgi.
- Une fois que le PI4P est transporté vers le réticulum endoplasmique (RE), son hydrolyse irréversible en PI par la phosphatase Sac1 associée au RE définit la directionnalité du cycle, permettant le transport du cholestérol depuis le RE pour enrichir le Golgi.
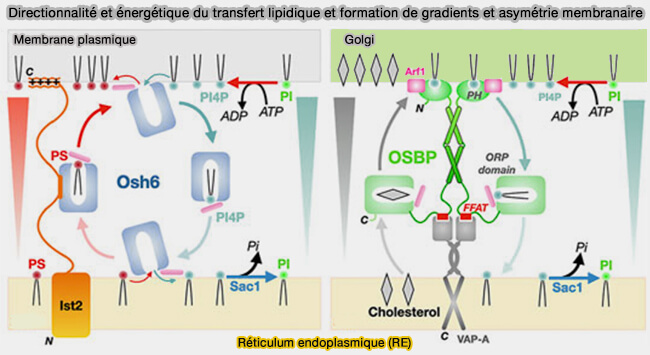
formation de gradients et asymétrie membranaire
(Figure : vetopsy.fr d'après Egea)
Étapes limitante du transfert lipidique
LTP en forme de panier
Dans le cas des LTP en forme de panier, l'étape limitante du transfert d'un lipide d'une membrane à une autre est l'extraction ou la désorption de la membrane elle-même plutôt que le transport assisté par LTP (Speed limits for non-vesicular intracellular sterol transport 2017 et Membrane hydrophobicity determines the activation free energy of passive lipid transport 2021).
- La désorption spontanée d’un lipide dans la phase aqueuse entre deux membranes est coûteuse en énergie (∼10–20 kCal/mol) et dure plusieurs jours.
- La composition locale de la membrane, l'emballage lipidique et les membranes en elles-mêmes affectent également l'énergie de la désorption lipidique.
1. Les LTP abaissent la barrière énergétique de désorption en séparant le lipide d'une bicouche dans une poche de liaison hydrophobe au lieu d'un solvant aqueux en vrac par deux mécanismes :
- l’électrostatique et la désolvatation/solvatation du site de liaison aux lipides (An electrostatic switching mechanism to control the lipid transfer activity of Osh6p 2019),
- les propriétés de remodelage ou de flexion membranaire des LTP cruciales pour la désorption au niveau de la membrane donneuse.
2. Les courbures observées dans le dimère E-SYT2 et le tétramère Mdm12-Mmm1 du complexe ERMES pourraient moduler la forme de la membrane (et/ou vice versa), réduisant ainsi l'énergie d'activation associée à la désorption et au transfert des lipides vers le noyau LTP.
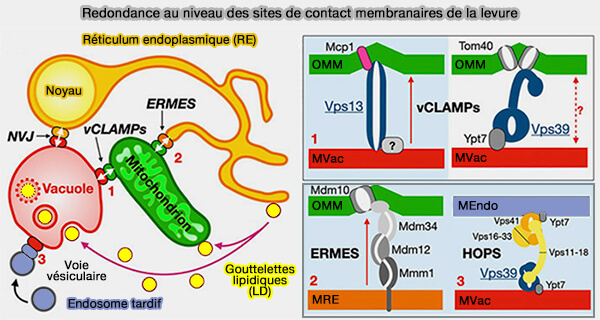
(Figure : vetopsy.fr d'après Egea)
2. Toutefois, ces mécanismes sont encore trop lents pour soutenir les fonctions cellulaires, i.e. quelques lipides par minute et d’autres facteurs, soit inconnus, soit difficilement mesurables/quantifiables dans les conditions expérimentales utilisées in vitro ou in vivo, doivent être pris en compte.
a. D'autres protéines pourraient augmenter la vitesse de désorption des lipides en déformant/amincissant localement la membrane comme :
- Mcp1 dans le cas de Vps13 (Vps13-Mcp1 interact at vacuole–mitochondria interfaces and bypass ER–mitochondria contact sites 2017),
- Mdm10 dans le cas d'ERMES,
- TOM40 avec Vps39.
L'importance de la distorsion et de l'amincissement de la membrane induits par les protéines transmembranaires, i.e. à base d'hélice α ou de brins β, a été documentée dans différents systèmes, en particulier les systèmes de translocation de protéines (Translocation of proteins through a distorted lipid bilayer 2021 et Highlighting membrane protein structure and function: A celebration of the Protein Data Bank 2021).
b. la redondance des sites de contact membranaire (MCS) jouerait aussi un rôle dans la vitesse de transfert lipidique.
LTP en forme de pont/tunnel
Quant aux transferts lipidiques par ponts/tunnels, le processus est beaucoup plus rapide car ils minimisent les effets de diffusion des protéines, et les seuls facteurs limitants apparents sont :
- la dynamique des lipides dans le tunnel protéique,
- la désorption des lipides.
Transfert lipidique par navette
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesGlycérophospholipidesSphingophospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation