Les P-ATPases, ou ATPases de type P, sont un grand groupe de pompes ioniques et lipidiques que l'on trouve dans les bactéries, les archées et les eucaryotes.
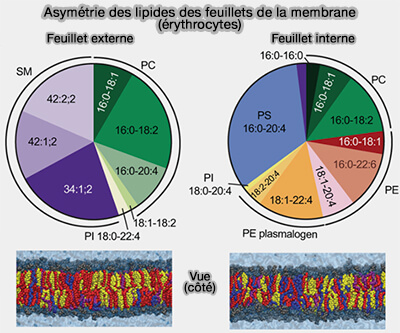
Asymétrie des lipides des feuillets membranaires
(Figure : vetopsy.fr d'après Lorent et coll)
Les flippases proprement dites déplacent les lipides de l'extérieur des membranes vers l'intérieur, i.e. milieu extracellulaire vers cytosol, ou cytosol vers le lumen des organites.
Ce processus génère et maintient une asymétrie lipidique entre les deux couches membranaires ( vue d'ensemble des flippases).
2. Ce processus, dans le cas des P4-ATPases comme dans celui des transporteurs ABC, la deuxième grande famille de flippases, fait appel à l'ATP.
Les lipides qui sont synthétisés dans les cellules sont incorporés dans des membranes au sein du feuillet interne, d'où des flippases peuvent les transférer dans le feuillet externe moyennant l'hydrolyse d'une molécule d'ATP.
Remarque : le cholestérol, un lipide majeur de la membrane plasmique, peut se déplacer librement entre les deux feuillets, mais est susceptible d'être plus concentré sur le feuillet exoplasmique en raison de son affinité pour la sphingomyéline (SM) et les glycosphingolipides.
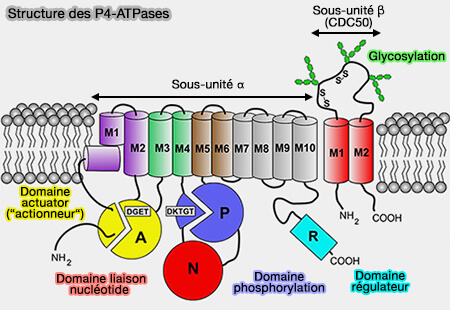
Les P4-ATPases possèdent une sous-unité α comme les autres P-ATPases avec en plus, une sous-unité β.
Les P-ATPases et leur structure sont étudiées dans un chapitre spécifique.
Structure du complexe Drs2p/Cdc50p
(Figure : vetopsy.fr d'après Timcenko et coll)
La P4-ATPase la plus étudiée actuellement est Drs2p, sous-unité α d'une flippase de la levure nécessaire au bourgeonnement des vésicules de transport à partir du réseau trans-Golgi (TGN) qui a pour sous-unité β Cdc50p.
Sous-unité α
La sous-unité α comprend plusieurs domaines.
1. Le domaine transmembranaire (M) possède un noyau central à 10 hélices α (M1-M10), i.e. M1-M6 étant commun à toutes les P-ATPases.
L'extension cytoplasmique C-terminale de la sous-unité α contient un domaine régulateur (R) ou autoinhibiteur.
2. Les domaines cytoplasmiques comprennent les trois domaines classiques des P-ATPases, impliqués dans le cycle catalytique de l'ATP similaires à ceux de la Ca++-ATPase et de Na+/K+-ATPases.
Le domaine de phosphorylation (P) contient le résidu d'acide aspartique (D) dans le motif DKTGT conservé qui subit une phosphorylation transitoire.
Le domaine actuator (A), actionneur en français, i.e. qui transforme l’énergie qui lui est fournie en un phénomène physique, contient le motif DGET (TGES dans Ca++-ATPase et Na+/K+-ATPase) qui facilite la déphosphorylation de l'intermédiaire phosphorylé.
Sous-unités β
1. La sous-unité β, i.e. protéine CDC50, comprend deux hélices transmembranaires dont l'ectodomaine se replie en deux lobes asymétriques, i.e. elle est absente dans ATP9A et 9B.
b. Le pli est similaire à celui de la seipine, protéine de liaison lipidique, bien que les boucles de Cdc50p soient considérablement plus longues.
3. Les deux hélices TM1 et TM2 de Cdc50p interagissant étroitement l'une avec l'autre et avec TM10 de la sous-unité α.
4. En l'absence d'expression de CDC50, les P4-ATPases ne parviennent pas à sortir du réticulum endoplasmique (RE), i.e. elles sont mal pliées, et sont incapables de subir une phosphorylation ou une hydrolyse de l'ATP dépendante des phospholipides.
Néanmoins, l'étude de Drs2P/Cdc50p a permis d'éclaircir un peu son rôle.
1. Du côté luminal de Drs2p, les interactions sont les plus étendues, i.e. l'ectodomaine de Cdc50p interagit avec toutes les boucles luminales en dehors de la boucle TM1-2.
Interactions Drs2p/Cdc50p
(Figure : vetopsy.fr d'après Timcenko et coll)
La boucle TM3-4 en particulier s'étend dans un site d'interaction à l'ectodomaine de Cdc50p.
2. Les contacts des régions transmembranaires semblent être moins nombreux, car seul TM10 de Drs2p interagit avec les hélices de Cdc50p.
3. Du côté cytosolique, l'extrémité N-terminale de Cdc50p s'enroule autour du domaine TM de Drs2p et établit des contacts avec le segment (résidus 529-538) qui relie TM4 et le site de phosphorylation du domaine P.
Ce segment est plus long de 10 résidus dans les P4-ATPases que dans les P2-ATPase, où TM4 couple le site de phosphorylation avec les conformations conformationnelles du domaine TM.
Le segment TM4 ainsi que la boucle TM3-4 sont nettement plus courts dans les flippases lipidiques Neo1p et homologues qui ne se lient pas aux sous-unités Cdc50p.
Ces interactions suggèrent que Cdc50p est sensible :
à l'état de phosphorylation du domaine P du côté cytosolique,
à la détection de la conformation du domaine transmembranaire par le contact avec la boucle TM3-41 et autres boucles du côté luminal.





 vue d'ensemble des flippases).
vue d'ensemble des flippases).