


Transport des lipides
Lipoprotéines : récepteurs
Récepteurs LDLR : LRP6 et LRP5 (structure et signalisations)
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Vue d'ensemble des lipides
- Acides gras
- Classification des lipides
- Cholestérol
- Glycérides
- Trafic non vésiculaire des lipides et LTP
- Digestion et absorption entérocytaire des lipides
- Lipoprotéines
- Gouttelettes lipidiques
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

LRP6 (LDL Receptor–Related Protein-6) et LRP5, membres éloignés de la famille des LDLR (Low-Density Lipoprotein Receptor), sont principalement des récepteurs de la voie de signalisation canonique Wnt.
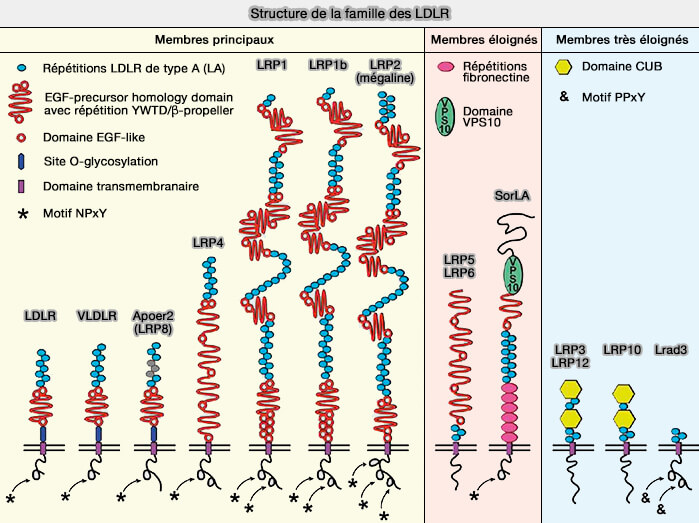
(Figure : vetopsy.fr d'après Pohlkamp et coll)
Les protéines de la famille LDLR partagent des motifs structurellement communs.
Structure de LRP5/6
Lrp5 et Lrp6 partagent 71 % d'homologie et sont des membres plus éloignés de la famille des LDLR.
1. La structure commune de LRP5/6, bien que présentant les mêmes domaines que des autres membres de la famille, est un peu différente (Crystal structures of the extracellular domain of LRP6 and its complex with DKK1 2011).
a. Lrp5 et Lrp6 possèdent :
- quatre β-propeller à six lames correspodant à six répétitions YWTD qui ne présentent que peu d'homologie entre elles, 19% chez l'homme,
- quatre domaines EGF-like, chacun suivant le β-propeller qui peut couvrir la face inférieure de l'hélice YWTD, comme pour LDLR,
- trois répétitions LDLR de type A (LA), appelé aussi LBR (Ligang Binding Repeat) ou LR qui, contrairement aux autres membres des LDLR, sont adjacents au domaine transmembranaire TM plutôt qu'à l'extrémité N-terminale.
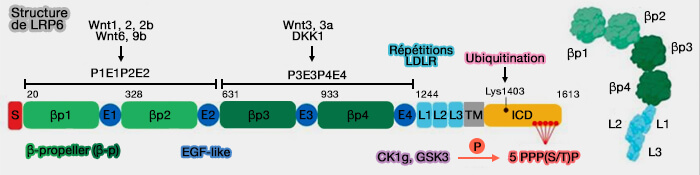
(Figure : vetopsy.fr d'après Raisch et coll)
b. leur domaine intracytoplasmique (ICD) ne possèdent pas de motif NPxY, mais cinq motif PPPS/TP (Pro-Pro-Pro-Ser/Thr-Pro) dans lesquels la sérine/thréonine peuvent être phosphorylées par l'activation Wnt (A Mechanism for Wnt Coreceptor Activation 2004).
Les kinases principales qui peuvent phosphoryler l'ICD sont (A dual-kinase mechanism for Wnt coreceptor phosphorylation and activation 2005 et Multiple PPPS/TP motifs act in a combinatorial fashion to transduce Wnt Signaling through LRP6 2009) :
- GSK-3 (Glycogen Synthase Kinase-3) pour les motifs PPPS/TP,
- CK1 (Casein Kinase1) pour les régions attenantes.
2. Les protéines Dishevelled (DVL) sont essentielles pour l'agrégation des LRP6 induite par Wnt avec Frizzled (FZD), et le complexe ternaire formé par LRP6, FZD et DVL repose sur ( activation des DVL) :
activation des DVL) :
-
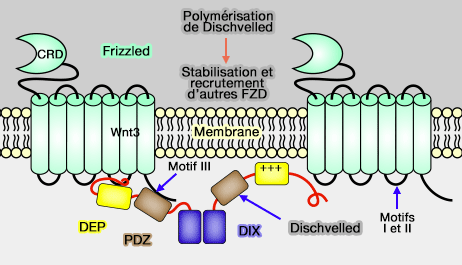
Polymérisation de Dischwelled et stabilsation
et recrutement de Frizzled
(Figure : vetopsy.fr d'après Tauriello) - les domaines PDZ de DVL qui se lient à Frizzeld.
Signalisation Wnt
La voie de signalisation Wnt est une voie essentielle comme dans l'embryogenène et la régénération cellulaire (Lrp5 and Lrp6 in Development and Disease 2013) et le cancer (Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities 2017).

Vous pouvez lire : Regulation of the Low-Density Lipoprotein Receptor-Related Protein LRP6 and Its Association With Disease: Wnt/β-Catenin Signaling and Beyond (2021).
Vue d'ensemble
Les voies Wnt régulent la différenciation et la spécialisation cellulaires en impliquent la liaison des molécules Wnt aux récepteurs Frizzled transmembranaires, via des mécanismes de signalisation distincts (WNT Signaling in Disease 2019) :
- la stabilisation " canonique " de la β-caténine régulant l'expression des gènes qui nécessite une interaction avec LRP5/6 (Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities 2017).
- les voies " non canoniques " de polarité cellulaire planaire (PCP) et Ca++/Calmoduline qui influencent le réarrangement du cytosquelette.

La voie de signalisation Wnt est étudiée dans un chapitre spécifique.
En l'absence de signal Wnt
1. En absence de signal Wnt, LR65/6 et le récepteur Frizzled de Wnt sont inactifs.
- Le pool cytoplasmique de β-caténine est recruté dans un complexe de destruction multiprotéique qui comprend l'axine, l'APC, CK1 (Casein Kinase1) et GSK-3 (Glycogen Synthase Kinase-3).
- La phosphorylation intracellulaire de la β-caténine est activée.
- Après phosphorylation, la β-caténine se combine avec le β-TRCP, une F-box de SCF, ce qui entraîne son ubiquitination et sa dégradation protéasomale ( dégradation de la β-caténine).
2. La disponibilité de LRP5/6 pour la signalisation est régulée par plusieurs mécanismes.
a. Des inhibiteurs sécrétés peuvent se lier directement à LRP5/6 et empêcher l'activation par les ligands Wnt comme :
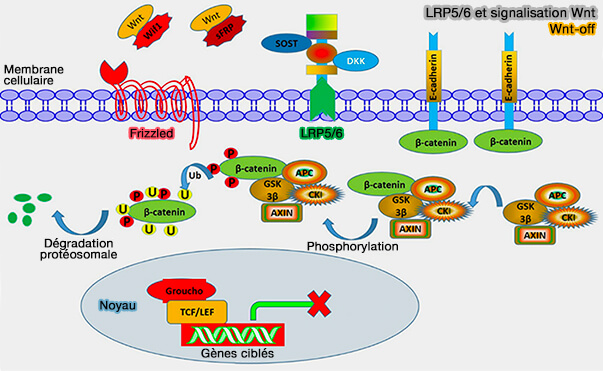
(Figure : vetopsy.fr d'après Gao et coll)
b. L'E3 ubiquitine ligase transmembranaire ZNRF3 diminue normalement la stabilité du complexe LRP5/6/Frizzled en ciblant le complexe pour la dégradation (ZNRF3 promotes Wnt receptor turnover in an R-spondin-sensitive manner 2012).
- Cependant, en présence de protéines R-spondine (Rspo), un complexe se forme entre ZNRF3, R-spondine (Rspo) et le récepteur LGR4.
- Ce mécanise provoque la clairance membranaire de ZNRF3, i.e. l'augmentation de la disponibilité du complexe LRP5/6/Frizzled potentialise la signalisation.
En présence d'un signal Wnt
L'activation de LRP5/6 peut se produire par plusieurs mécanismes. La plus connue se produit lorsqu'un ligand Wnt engage un récepteur Frizzled, et LRP5/P6.
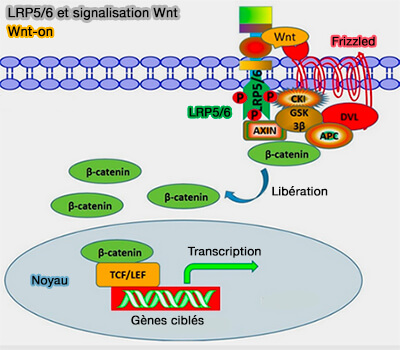
(Figure : vetopsy.fr d'après Gao et coll)
1. En présence d'un signal Wnt, i.e. Wnt-on, le ligand Wnt se lie au récepteur Frizzled lié à Dischevelled (DVL) et au co-récepteur LRP5/6 à la surface de la membrane cellulaire.
- L'activation du complexe récepteur Wnt entraîne la phosphorylation par CK1 et GSK-3β de Thr1479 et Ser1490 dans l'extrémité C-terminale de LRP6 au niveau du motif PPPS/TP (Wnt3a-mediated formation of phosphatidylinositol 4,5-bisphosphate regulates LRP6 phosphorylation 2008).
- Ce processus crée un site de liaison pour l'axine, la séquestrant dans le complexe de signalisation et l'empêchant de contribuer au complexe de destruction de la β-caténine.
- Il en résulte une accumulation de la β-caténine et sa translocation dans le noyau, où elle peut se complexer avec les membres de la famille LEF/TCF pour réguler la transcription des gènes cibles (β-caténine et activation génique).
2. Un autre acteur important du signalosome est PIP5K1, une phosphatidylinositol phosphate kinase dont l'activation est médiée par FZD et DVL (Wnt3a-mediated formation of phosphatidylinositol 4,5-bisphosphate regulates LRP6 phosphorylation 2008).
-
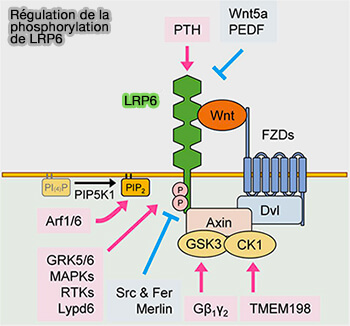
Régulation de la phosphorylation de LRP6
(Figure : vetopsy.fr d'après Jeong et Jho) - Par conséquent, les composants non protéiques tels que les phospholipides (PL) peuvent également jouer un rôle crucial dans la signalisation Wnt/β-caténine médiée par LRP6.
Remarque : d'autres kinases peuvent intervenir dans la phosphorylation de LRP5/6 et entrer dans la régulation.
3. La norrine peut également activer la signalisation β-caténine de manière dépendante de Frizzled-4 et LRP5 ( activation de la voie Wnt par la norrine).
4. Le complément C1q peut se lier à Frizzled et activer les C1 pour cliver LRP5/6, créant une version tronquée de cette protéine qui est constitutivement active (Complement C1q Activates Canonical Wnt Signaling and Promotes Aging-Related Phenotypes 2012).
- Les taux sériques de C1q augmentent avec l'âge.
- La signalisation Wnt dépendante de C1q altère la capacité de régénération des muscles squelettiques.
Internalisation de LRP6
L'internalisation des récepteurs, après la liaison aux ligands, joue un rôle crucial dans la transduction du signal en régulant positivement ou négativement la voie de signalisation canonique Wnt.
Pour une revue lire : Regulation of the Low-Density Lipoprotein Receptor-Related Protein LRP6 and Its Association With Disease: Wnt/β-Catenin Signaling and Beyond (2021).
1. L'exemple le plus connu est DKK1 (Dickkopf1) qui forme un complexe ternaire avec Kremen, la protéine transmembranaire ( régulation par Kremen).
- Ce complexe internalise LRP6 et diminue ses niveaux de membrane plasmique, conduisant à l'inhibition de la signalisation Wnt/β-caténine .
- L'internalisation de LRP6 via Dkk1 entraîne également une diminution de la phosphorylation de LRP6 par CK1.
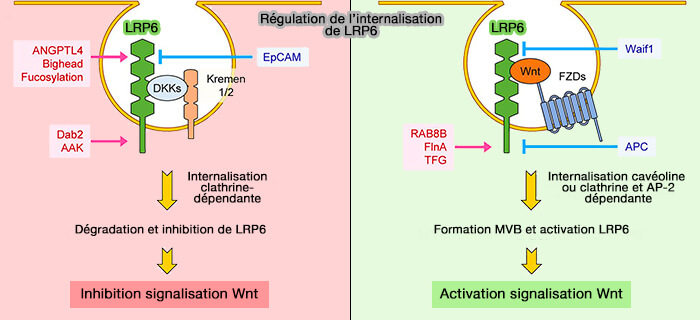
(Figure : vetopsy.fr d'après Jeong et Jho)
2. D'autres régulateurs peuvent intervenir :
- ANGPTL4 (ANGioProTéines-Like 4), forme un un complexe avec les syndécanes et LRP6 et induit l'internalisation de LRP6 et diminue ses niveaux de surface (Angiopoietin-like 4 Is a Wnt Signaling Antagonist that Promotes LRP6 Turnover 2017).
- la fucosylation de LRP6 favorise l'internalisation de la LRP6 localisée dans le radeau lipidique et inhibe la formation du complexe Wnt-FZD-LRP6 et inhibe ainsi la signalisation Wnt/β-caténine (In Situ Fucosylation of the Wnt Co-receptor LRP6 Increases Its Endocytosis and Reduces Wnt/β-Catenin Signaling, 2020).
- Waif1 (Wnt-activated inhibitory factor 1) ou 5T4, une protéine transmembranaire, interagit avec LRP6 et inhibe l'internalisation de LRP6 induite par Wnt3a dans les vésicules endocytaires, réduisant ainsi la signalisation Wnt/β-caténine (Waif1/5T4 Inhibits Wnt/β-Catenin Signaling and Activates Noncanonical Wnt Pathways by Modifying LRP6 Subcellular Localization 2011),
- …
Autres signalisations
LRP6 affecte non seulement la signalisation Wnt/β-caténine, mais également d'autres voies de signalisation comme :
- les GPCR (récepteurs couplés aux protéines G),
- la voie de signalisation Wnt non canonique, i.e. ROR1/2,
- la voie de signalisation Hippo (LDL receptor-related protein LRP6 senses nutrient levels and regulates Hippo signaling 2020),
- la signalisation de stabilisation des protéines dépendante de Wnt (Wnt/STOP).
Pour une revue lire : Regulation of the Low-Density Lipoprotein Receptor-Related Protein LRP6 and Its Association With Disease: Wnt/β-Catenin Signaling and Beyond (2021).
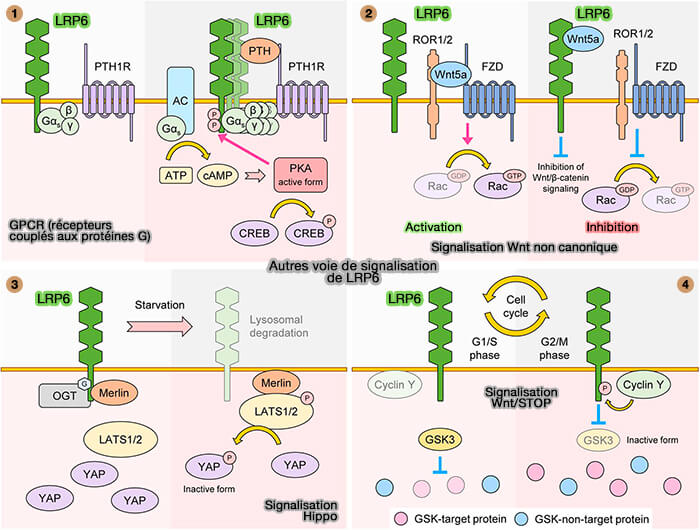
(Figure : vetopsy.fr d'après Jeong et Jho)
Fonctions de LRP5/6
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesFormation des chylomicronsHDL et remodelageTrafic des lipoprotéinesRécepteurs des lipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation