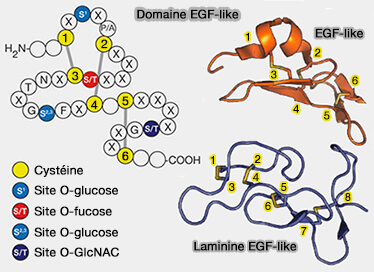
1. Les domaines du type EGF sont présents dans quelques centaines de protéines de surface cellulaire, i.e. protéines membranaires ou dans les protéines connues pour être sécrétées.
au développement neuronal par la famille Notch de récepteurs et les ligands canoniques Notch (Delta et Serrate/Jagged) impliqués dans les décisions du destin cellulaire,
l'adhésion cellulaire,
la reconnaissance des protéines pour leur ciblage ou leur dégradation.
b. Ces domaines sont souvent impliqués dans la libération des ligands des endosomes, mais aussi dans les liaisons avec d'autres protéines.
Domaine EGF-like
(Figure : vetopsy.fr d'après Varki et Tombling)
dans de grandes protéines qui peuvent être O-glycosylées comme la fibrilline, glycoprotéine essentielle à la formation des fibres élastiques du tissu conjonctif à plus de 30 domaines EGF, ou les laminines, famille de glycoprotéines de la matrice extracellulaire (ECM) des animaux,
Les domaines EGF se retrouvent aussi en tandem dans des protéines dans lesquelles leur repliement forme un domaine solénoïde linéaire.
On le retrouve souvent avec des répétitions d'autres modules comme le domaine Kringle, intervenant dans les interactions protéine-protéine avec les facteurs de coagulation sanguine ou les Lp(a).
Structure du domaine EGF-like
Le domaine EGF-like, d'une quarantaine de résidus, est variable selon les espèces (60 à 70% d'homologie).
1. Les modifications des glycanes des protéines à domaine EGF régulent la transduction du signal pendant :
le développement embryonnaire et le maintien des organes adultes,
la différenciation cellulaire,
la croissance de plusieurs cancers.
Les mutations de plusieurs enzymes impliquées dans l'addition ou l'élongation de ces glycanes ont été retrouvées dans des maladies humaines (Congenital Disorders of Glycosylation 2022).
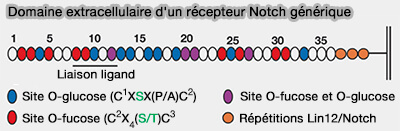
2. Un bon exemple est le domaine extracellulaire de Notch qui contient jusqu'à 36 répétitions EGF en tandem, dont un grand nombre contiennent des sites consensus pour un ou plusieurs types d'O-glycane (Other Classes of Eukaryotic Glycans 2022).
O-glycosylations du récepteur Notch
(Figure : vetopsy.fr d'après Varki et coll)
Les répétitions Notch EGF peuvent héberger des O-fucoses, des O-GlcNAc et/ou des O-glucoses.
Ces modifications de sucre régulent divers aspects de la transduction du signal Notch, parfois de manière partiellement redondante et parfois via des mécanismes distincts.
3. Les mutations de toutes les enzymes de ces O-glycosylations provoquent des maladies génétiques.
Glycanes O-fucose
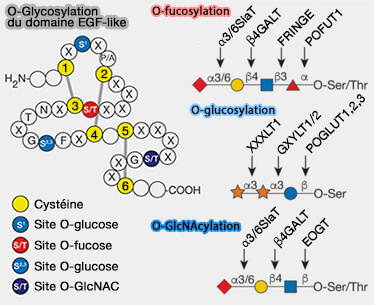
1. Cette O-fucosylation survient sur la sérine/thréonine avant la troisième cystéine et est trouvée dans plus d'une centaine de protéines.
L'O-fucose peut être allongé en un tétrasaccharide, i.e. Siaα2–3/6Galβ1-4GlcNAcβ1-3Fucα-O-Ser/Thr, comme dans l'EGF1 du facteur IX de coagulation humain.
2. Les glycanes O-fucosylés par POFUT1 régulent principalement, sinon exclusivement, la signalisation Notch en ajoutant des glycanes O-fucose aux récepteurs Notch et participent à la liaison canonique du ligand Notch.
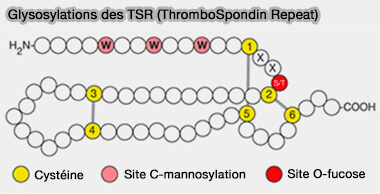
Glycosylations des TSR (ThromboSpondin Repeat)
(Figure : vetopsy.fr d'après d'après Varki et coll)
1. Cette O-glucosylation survient sur la sérine entre la première et la seconde cystéine, et est trouvée dans plus d'une cinquantaine de protéines.
Le motif consensus est $\ce{C^1XSX(P/A)C^2}$.
L'O-glucosylation est réalisée par la O-glucosyltransférase 1 (POGLUT1).
Le glycane O-glucose existe généralement sous la forme du trisaccharide, i.e. Xylα1–3Xylα1–3Glcβ-O-Ser.
2. Récemment, un deuxième site de modification de l'O-glucose lié en β a été identifié entre la troisième et la quatrième cystéine des répétitions EGF et est trouvé dans plus d'une trentaine de protéines.
Le motif consensus supposé est $\ce{C^3XNTXGSFXC^4}$.
Les homologues de POGLUT1, appelés POGLUT2 et POGLUT3, sont responsables de la modification de ce site.
L'O-glucose à ce site n'a pas été signalé comme étant allongé au-delà du monosaccharide.
Glycanes O-GlcNAc
Bien que l'O-GlcNAc liée à la β soit très courante sur les protéines des compartiments nucléaire et cytosolique, la même modification peut être trouvée dans relativement peu de protéines membranaires et sécrétées.
Cette O-GlcNAcylation, i.e. ajout de N-acétylglucosamine, survient sur la sérine/thréonine entre la cinquième et la sixième cystéine et est trouvée dans plus d'une cinquantaine de protéines.
Le motif consensus est $\ce{C^5XXXX(S/T)GX_{2-3}C^6}$.




 cf. tableau).
cf. tableau).