


Trafic vésiculaire :
Endocytose clathrine-indépendante (CIE)
Cavéoles
Biogenèse
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Transports membranaires sans mouvements membranaires
- Transports membranaires avec mouvements membranaires : trafic vésiculaire
- Protéines membranaires
- Moteurs moléculaires
- Voies de signalisation

La biogenèse des cavéoles est un phénomène complexe qui implique l'action coordonnée de plusieurs protéines et des lipides membranaires.
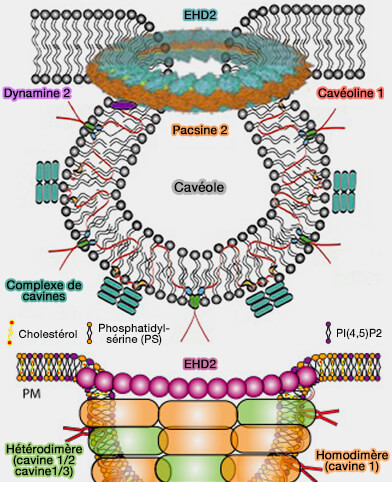
(Figure : vetopsy.fr d'après Lian et coll et Parton et coll)
1. La formation de la fosse cavéolaire implique l'action coordonnée de plusieurs protéines (Caveolae 2018).
a. Les protéines membranaires intégrales, i.e. les cavéolines (CAV), sont essentielles à leur formation, i.e. leur KO inhibe la genèse cavéolaire.
b. Les protéines de la membrane périphérique, i.e. les cavines, interagissent avec les cavéolines pour former un complexe de revêtement externe qui remodèle la membrane plasmique (MP) en cavéoles et les stabilisent par leurs interactions avec les lipides membranaires, i.e. cholestérol, phosphatidylsérine (PS) et PI(4,5)P2 ou PIP2.
c. Les complexes protéiques au niveau du cou impliquent :
- des protéines EHD comme EHD2 (Eps-15 Homology Domain 2), une ATPase formant un anneau autour du cou cavéolaire,
- les syndapines/PACSINes, i.e. syndapine 2/3 des protéines à domaine F-BAR, interviendraient (parfois ou toujours ?) pour remodeler la membrane.
Remarque : des protéines transmembranaires comme ROR1 (Receptor tyrosine kinase-like Orphan Receptor 1), qui interagit avec CAV1 et la cavine1 pourraient faciliter la formation de cavéoles, mais pas dans tous les tissus.
2. La composition lipidique de nanodomaines membranaires, en particulier en cholestérol et en phospholipides chargés négativement, i.e. phosphatidylsérine (PS) et PI(4,5)P2 ou PIP2, est indispensable à la biogenèse des cavéoles.
L'assemblage ainsi que les interactions entre toutes ces molécules, i.e. protéines/lipides, protéines/protéines et lipides/lipides sont loin d'être connus, ce qui implique que certains articles un peu anciens ne sont plus d'actualité et que les modèles sont nombreux.
Biogenèse moléculaire
1. La biogenèse des cavéoles s'effectue tout au long d'un processus complexe (Biogenesis of Caveolae: Stepwise Assembly of Large Caveolin and Cavin Complexes 2010).

Ce chapitre est très largement inspiré de : Caveolins and cavins in the trafficking, maturation, and degradation of caveolae: implications for cell physiology (2017).
Les cavéolines (Cav) étant les principaux composants protéiques des cavéoles, influençant la composition de la membrane et la teneur en protéines de la traduction à la dégradation, leur traduction et leur oligomérisation précoce commencent la voie de biogenèse des cavéoles qui s'effectue par étapes.
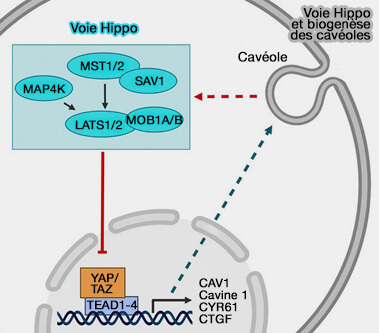
(Figure : vetopsy.fr d'après Lian et coll et Parton et coll)
Remarque : la voie de signalisation Hippo YAP/TAZ est critique pour l'expression de la cavine 1 et de la cavéoline-1, et donc pour la formation de cavéoles (The Hippo Pathway Regulates Caveolae Expression and Mediates Flow Response via Caveolae 2019).
- La voie consiste en une série de kinases qui peuvent contrôler la localisation subcellulaire et la stabilité de YAP/TAZ, i.e. Yes-Associated Protein/Transcriptional co-Activator with PDZ-binding motif, des cofacteurs transcriptionnels homologues.
- Les gènes cavéolaires essentiels CAVIN1 et CAVEOLIN1 sont des gènes cibles directs YAP/TAZ-TEAD.
- YAP/TAZ sont hyperactivés dans les cellules déficientes en cavéoles.
- Les cavéoles facilitent la réponse au stress de cisaillement médiée par YAP/TAZ.
1. L'insertion co-traductionnelle des monomères de Cav s'effectue dans la membrane du réticulum endoplasmique (RE) d'une manière dépendante des particules de reconnaissance du signal (SRP).
2. Les Cav s'oligomérisent en 8S-Cav, i.e. 7-14 Cav dans un rapport Cav1:Cav2 de 2–4:1 (Molecular composition and ultrastructure of the caveolar coat complex 2013).
La formation d'oligomères 8S-Cav dépend des résidus situés dans l'OD (Oligomerization Domain), i.e. dans le domaine SM (66–70 dans Cav1), dans le domaine SD (181–100) et dans la SR (Spoke region) 134–178.
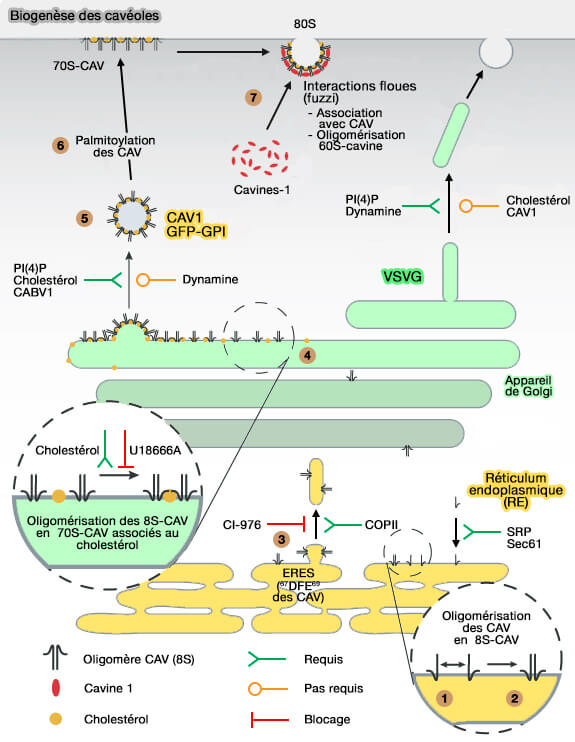
(Figure : vetopsy.fr modifié d'après Hayer et coll)
3. Le motif diacide (DXE, i.e. 67DFE69 dans Cav1) proche du SM permet une accumulation rapide dans l'ERES (ER Exit Site) et la sortie des Cav par les vésicules COPII.
Une forme mutante de la cavéoline-1 associée à l'hypertension artérielle pulmonaire s'accumule dans le RE en raison d'une altération de l'extrémité C-terminale contenant un motif de rétention RE, i.e. 176KKYK179 (A disease-associated frameshift mutation in caveolin-1 disrupts caveolae formation and function through introduction of a de novo ER retention signal 2017).
4. Dans l'appareil de Golgi, les oligomères 8S-Cav s'oligomérisent en 70S-Cav, i.e. 160 Cav1 et Cav2 dans 15–25 8S-Cav, qui s'associent avec le cholestérol par le motif CRAC, 94VTKYWFYR101 (Cholesterol and Fatty Acids Regulate Dynamic Caveolin Trafficking through the Golgi Complex and between the Cell Surface and Lipid Bodies 2005).
- Cette voie de sécrétion précoce est notamment sensible aux perturbations lipidiques (Conserved Functions of Ether Lipids and Sphingolipids in the Early Secretory Pathway 2020).
- En effet, une expression élevée de la cavéoline peut être nocive (
 ubiquitination des cavéolines).
ubiquitination des cavéolines).
5. Les oligomères 70S-Cav sont exportés du Golgi vers la membrane plasmique (MP) dans des vésicules sécrétoires indépendantes de la dynamine.
- FAPP-1 et FAPP-2 (Four-phosphate-adaptor-protein-1 et 2), protéines adaptatrices, interagissent avec PI(4)P pour générer des vésicules de transport (FAPPs control Golgi-to-cell-surface membrane traffic by binding to ARF and PtdIns(4)P 2004).
- Ces petites vésicules uniformes de 70S-Cav-cholestérol peuvent aussi transporter des cargos, comme des protéines liées au glycosylphosphatidyl inositol (GPI), environ 150 protéines chez l'homme, qui s'associent aux radeaux lipidiques (Intracellular Retention of Glycosylphosphatidyl Inositol-Linked Proteins in Caveolin-Deficient Cells 2002 et Biosynthesis and biology of mammalian GPI-anchored proteins 2020).
6. Les oligomères 70S-Cav subissent une palmitoylation près de la MP (C133, C143, C156 de Cav1) dont le rôle n'est pas clair car elle n'est pas indispensable à la liaison membranaire ( palmitoylation des cavéolines).
(Figure : vetopsy.fr modifié d'après Hayer et coll)
7. Lors de l'arrivée à la membrane plasmique (MP), 60-80 cavines s'oligomérisent (60S-Cavin) en rejoignant les oligomères 70S-Cav et les membranes riches en cholestérol et forment les stries caractéristiques des cavéoles (Architecture of the caveolar coat complex 2016 et Structural Insights into the Organization of the Cavin Membrane Coat Complex 2014).
- Les cavines s'oligomérisent en trimère de trimère (60S-Cavin) en l'absence de Cav1, mais elles sont progressivement ajoutées au 70S-Cav au PM pour former un complexe 80S ( oligomérisation des cavines).
- Les cavines ne se lient pas aux Cav au niveau de la membrane, excepté la cavine 3 (remarques sur les cavines).

L'oligomérisation des cavines, les interactions cavines/cavéolines font intervenir des interactions intramoléculaires dites " floues ", fuzzy en anglais ( rôles des DR des cavines).
(Figure : vetopsy.fr d'après Zhou et coll)
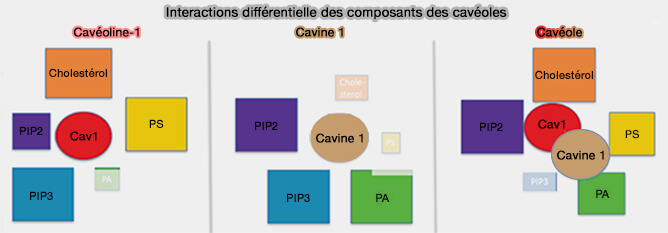
Les cavines ont des sites de liaison aux phospholipides anioniques, i.e phosphatydylsérine (PS) dans HR2 de la cavine 1 (domaine UC1) et PI(4,5)P2 ou PIP2 dans HR1 qui sont concentrés localement dans les membranes des cavéoles ( cavines et lipides membranaires).
La fusion des vésicules avec la PM, i.e. pré-invagination, permet aux protéines ancrées au GPI de diffuser latéralement tandis que la couche Cav reste au point d'intégration.
7. Lors de l'invagination membranaire, les cavéolines s'associent aux cavines dans des domaines discrets de la membrane plasmique pour former des cavéoles par des interactions floues d'affinité relativement faible, nécessitant une oligomérisation des protéines cavine et cavéoline ainsi que des interactions avec lipides membranaires.
- Les cavéolines interagissent avec les lipides membranaires dans la bicouche tandis que les cavines s'associent et regroupent des lipides membranaires spécifiques dans le feuillet cytoplasmique de la membrane plasmique (Caveolin-1 and cavin1 act synergistically to generate a unique lipid environment in caveolae 2021).
- Les multiples interactions entre les protéines cavéolaires et différents lipides membranaires, un processus appelé détection de coïncidence, sont cruciales pour former le domaine de la cavéole métastable (Coincidence detection in phosphoinositide signaling 2005).
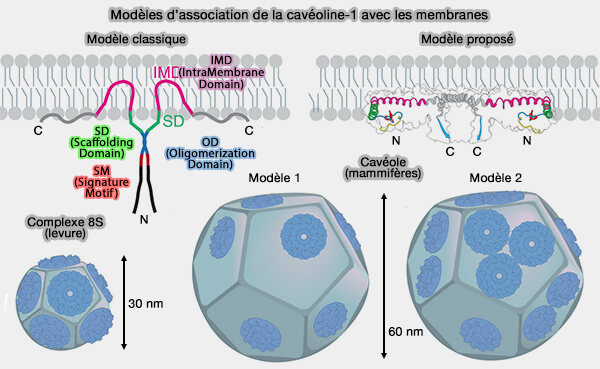
(Figure : vetopsy.fr d'après Porta et coll)
a. Les domaines IMD des Cav, selon le modèle dominant, forment une structure en épingle à cheveux qui s'insère dans la membrane, créant un coin qui plie la membrane (Key Phases in the Formation of Caveolae 2021). Un autre modèle peut être proposé (Molecular architecture of the human caveolin-1 complex 2022).
- De plus, les trois cystéines palmitoylées des Cav stabilisent les oligomères Cav dans la membrane des cavéoles et peuvent participer à la courbure de la membrane (Probing the U-Shaped Conformation of Caveolin-1 in a Bilayer 2014).
b. Les DR des cavines sont impliquées dans la courbure membranaire comme dans la liaison avec les cavéolines par les interactions floues ( DR et courbure membranaire).
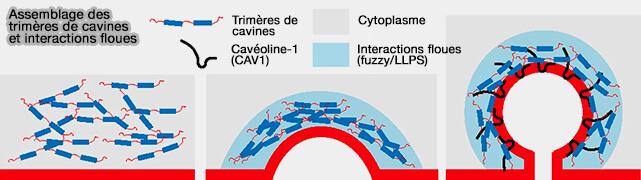
(Figure : vetopsy.fr d'après Tillu et coll)
Remarque : cependant, dans des conditions normales, le processus de génération de la courbure membranaire, pour restreindre l'activité de remodelage de la cavine 1 uniquement aux cavéoles. est aussi étroitement régulé par :
- EHD2 qui stabilise le cou des cavéoles et inhibe le processus de bourgeonnement favorisé par le cholestérol ( cavéoline et cholestérol),
- la syndapine 2 qui stabilise aussi le cou des cavéoles par son interaction avec EHD2 ( syndapine 2/PACSINe 2 et stabilité membranaire),
- Cav1 et sa liaison avec la filamine A ( cavines et cytosquelette),
- des lipides membranaires spécifiques, tels que PI(4)P dans les vésicules en provenance de l'appareil de Golgi, peuvent également contribuer à la courbure de la membrane et à l'invagination des cavéoles (Phosphatidic acid in membrane rearrangements 2019).
Résumé du modèle de biogenèse
Les modèles de formation des cavéoles peuvent différer. Nous utiliserons le modèle simplifié décrit dans : Key phases in the formation of caveolae (2021).
1. Les cavéolines atteignent le PM, arrivant du complexe de Golgi sous forme de structure d'oligomères riches en cholestérol pour générer des nanodomaines enrichis en un sous-ensemble de lipides membranaires (Super-resolution modularity analysis shows polyhedral caveolin-1 oligomers combine to form scaffolds and caveolae 2019).
La cavéoline serait, selon un des modèles, insérée partiellement dans la membrane ( insertion des cavéolines dans la membrane).
2. Ces domaines agissent comme des plates-formes pour recruter les oligomères cytoplasmiques de cavines aux cavéoles via des interactions floues entre domaines désordonnés, aidant à créer le domaine lipidique nécessaire à l'insertion complète de la protéine cavéoline dans la membrane par son interaction avec des lipides membranaires spécifiques (Caveolin-1 and cavin1 act synergistically to generate a unique lipid environment in caveolae 2021).
La mutation du domaine de liaison PI(4,5)P2 ou PIP2 de la cavine 1 provoque un changement non seulement dans l'association de PI(4,5)P2 avec les cavéoles, mais provoque des modifications de plusieurs lipides, y compris du cholestérol.
Les cavines, chez les mammifères, joueraient donc un rôle crucial dans la promotion de la formation d'un nanoenvironnement lipidique qui facilite l'interaction de la cavéoline avec la bicouche propice à la formation de cavéoles et qui régule la formation et le désassemblage des cavéoles.
3. Dans ce modèle, les cavines ne sont pas absolument essentielles pour générer un domaine de type cavéole : les systèmes membranaires avec une composition lipidique adéquate peuvent favoriser l'insertion de cavéoline et la formation de cavéoles en l'absence de cavines.
- D'une part, c'est ce qu'on observe dans des systèmes cellulaires plus primitifs, i.e. bactéries ou invertébrés (Constitutive Formation of Caveolae in a Bacterium 2012 et Ascidian caveolin induces membrane curvature and protects tissue integrity and morphology during embryogenesis 2020).
- D'autre part, cela expliquerait les observations selon lesquelles l'addition de lipides peut favoriser une courbure et une scission plus élevées de la cavéole et la dépendance de lipides spécifiques sur l'association cavine avec la cavéoline et la formation de cavéoles.
En outre, les formations tubulaires créées par l'activité membranaire des cavines pourraient contribuer à la formation d'un domaine cavéolaire très incurvé stable (Structural Insights into the Organization of the Cavin Membrane Coat Complex 2014 et Cavin1 intrinsically disordered domains are essential for fuzzy electrostatic interactions and caveola formation 2021).
4. Ce modèle est bien entendu simplifié, car d'autres molécules sont impliquées comme :
- ROR1,
- les syndapines/PACSINes dans les cellules musculaires sûrement, mais est-ce le cas dans les autres cellules ( KO pacsine 3) ?

Les cavéoles auraient évolué pour devenir des capteurs très sensibles de la composition lipidique de la membrane plasmique qui peut prendre deux formes.
- L'aplatissement des cavéoles libère des cavines dans le cytosol pour la signalisation en aval et potentiellement libère des lipides membranaires de la membrane pour d'autres interactions.
- La génération d'une courbure plus forte, la rétention de cavine 1 et la scission génèrent un porteur vésiculaire (EHD2-mediated restriction of caveolar dynamics regulates cellular fatty acid uptake 2020 et Lipid accumulation controls the balance between surface connection and scission of caveolae 2020).
Retour vers les cavéoles
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseEndocytose clathrine-dépendante (CME)Endocytoses clathrine-indépendantes (CIE)CavéolesFEMEVoie CLIC/GEECVoie flotillineVoie Arf6IL2RGFR-NCEADBEUFEPinocytoseMacropinocytosePhagocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation