Le complexe SCF, E3 ubiquitine ligase de type RING, déclenche la dégradation régulée des protéines clés de la transition G1/S du cycle cellulaire.
Vue d'ensemble du complexe SCF
Le complexe SCF (Skp1-Cullin1-F-box) est une ligase CRL (Cullin-RING ligases) assurant la reconnaissance sélective et l’ubiquitination de nombreux régulateurs cellulaires, impliquée dans la dégradation contrôlée des régulateurs du cycle cellulaire et de multiples effecteurs de signalisation.
Sa structure modulaire, i.e. Skp1, CUL1, Rbx1 et une protéine F-box, lui confère une remarquable plasticité fonctionnelle grâce à la diversité de ses sous-unités F-box, véritables modules de spécificité.
1. Dans le cycle cellulaire, le SCF intervient en fin de G1 et lors de la transition G1/S, où il élimine les inhibiteurs clés des CDK (Cyclin-Dependent Kinases) et plusieurs facteurs limitants de la réplication.
Il agit de manière complémentaire à l’APC/C, ce dernier contrôlant la sortie de mitose tandis que le SCF sécurise l’engagement vers la phase S.
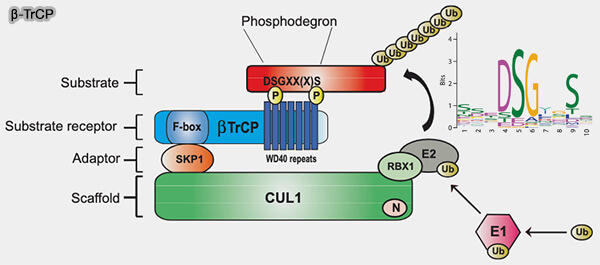
La capture des substrats repose sur la présence de phospho-dégrons, générés par les cyclines/CDK, CK1 ou GSK3β, garantissant un couplage direct entre signalisation mitogénique et décision de dégradation.
2. Au-delà du cycle, le SCF joue un rôle central dans de nombreuses voies comme ( substrats majeurs de SCF) :
la régulation de nombreuses voies de signalisation,
la réponse au stress,
le contrôle de la stabilité de multiples facteurs métaboliques
Le SCF doit être considéré comme un hub global, capable d’intégrer signaux externes, état métabolique et programmes transcriptionnels, tout en assurant un contrôle précis de la progression du cycle cellulaire.
la phosphorylation préalable du substrat, créant un phospho-dégron.
la condition de reconnaissance, transformation du segment phosphorylé en un motif tridimensionnel lisible par le domaine WD40, LRR ou un autre module de la F-box.
la reconnaissance spécifique par une protéine F-box adaptatrice, i.e. Skp2, β-TrCP, FBXW7… détermine la sélectivité du SCF en captant spécifiquement le motif phosphorylé,
le transfert catalytique d’ubiquitine via CUL1-Rbx1-E2~Ub, étape catalytique conduisant à l’ubiquitination.
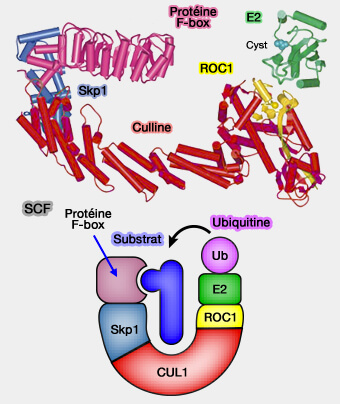
1. La culline1 (CUL1) forme le module d'échafaudage ou la colonne vertébrale du SCF.
a. Elle est composée par :
SCF
(Figure : vetopsy.fr)
un domaine N-terminal qui interagit avec Skp1,
une extrémité C-terminale (WHB) s’associe à Rbx1, qui contient le domaine RING responsable du recrutement de l’E2~Ub.
b. Cette configuration impose une distance et une orientation précises entre le substrat et l’E2~Ub, élément clé de l’efficacité catalytique.
2. Le module de reconnaissance est formé par deux protéines.
a. Skp1 (S-phase kinase-associated protein 1) stabilise l’interaction entre CUL1 et la F-box, créant une plateforme de capture spécifique.
b. Les F-box (FBP)se distinguent par leurs domaines de reconnaissance (WD40, LRR ou modules divers), qui déterminent la nature des phospho-dégrons et, par conséquent, l’identité des substrats recrutés.
La diversité de ces domaines explique la polyvalence fonctionnelle du SCF.
Les protéines F-box sont étudiées dans un chapitre spécifique.
3. Le module catalytique est constitué par Rbx1 et l’E2~Ub.
Rbx1 (RING-box protein 1), appelé aussi ROC1 (Regulator Of Cullins1) est doté d'un domaine RING qui recrute l’enzyme de conjugaison E2~Ub.
L’ancrage de Rbx1 à CUL1, associé à la flexibilité contrôlée du bras catalytique, permet d’aligner précisément l’E2~Ub avec les lysines accessibles du substrat ancré sur la F-box.
Cette géométrie conditionne à la fois l’initiation et l’allongement des chaînes d’ubiquitine.
SCFβ-TrCP
(Figure : vetopsy.fr d'après Low et coll)
Cette architecture spatiale positionne le substrat et l’enzyme de conjugaison dans une configuration catalytiquement optimale.




 substrats majeurs de SCF) :
substrats majeurs de SCF) :