Les cassures double brin de l’ADN constituent des lésions particulièrement dangereuses pour le génome et sont réparées par plusieurs voies dont la recombinaison homologue (HR), mécanisme fidèle utilisant la chromatide sœur comme matrice.
La réparation de l’ADN correspond spécifiquement à la correction enzymatique des lésions affectant la molécule d’ADN et repose sur plusieurs systèmes spécialisés :
a. réparant des lésions de l’ADN sans cassure double brin :
En l’absence de réparation rapide, ces lésions peuvent conduire à des délétions, des translocations chromosomiques ou à la perte de fragments chromosomiques, compromettant gravement la stabilité du génome.
1. Les cassures double brin peuvent apparaître :
a. de manière accidentelle, sous l'effet de différents mécanismes cellulaires ou environnementaux :
b. De manière programmée dans certains contextes biologiques où des réarrangements contrôlés de l'ADN sont nécessaires, notamment lors de différents mécanismes de recombinaison.
2. La recombinaison correspond à un ensemble de processus permettant l'échange, le réarrangement ou le transfert d'informations génétiques entre molécules d'ADN.
a. La recombinaison homologue erst impliquée dans la réparation de l'ADN et la méiose (recombinaison méiotique),
Les protéines RAG1 et RAG2 reconnaissent des séquences signal de recombinaison (RSS) situées de part et d’autre des segments géniques V (Variable), D (Diversity) et J (Joining).
Au cours de la maturation des lymphocytes, ces segments sont réarrangés de manière aléatoire pour former un gène fonctionnel, par exemple. V + J pour les chaînes légères des anticorps et certaines chaînes des récepteurs des lymphocytes T (TCR) ou V + D + J pour les chaînes lourdes des anticorps et les chaînes β des TCR.
Les protéines RAG1-RAG2 induisent alors des cassures double brin programmées qui permettent le réarrangement de ces segments par la voie NHEJ (Non-Homologous End Joining), générant un grand nombre de combinaisons possibles et contribuant ainsi à la diversité des récepteurs antigéniques du système immunitaire adaptatif.
c. La recombinaison spécifique de site repose sur la reconnaissance de courtes séquences d'ADN par des recombinases spécialisées qui catalysent l'insertion, l'excision ou l'inversion de segments d'ADN sans nécessiter de longues régions d'homologie.
Remarque : les mécanismes de transposition assurent le déplacement de séquences d'ADN appelées éléments transposables au sein du génome ( ADN répétitif).
Les transposons à ADN utilisent principalement des transposases, tandis que les rétrotransposons reposent sur des transcriptases inverses associées à des endonucléases ou des intégrases.
Bien qu'ils puissent provoquer des réarrangements génomiques, ils sont généralement distingués des mécanismes de recombinaison proprement dits.
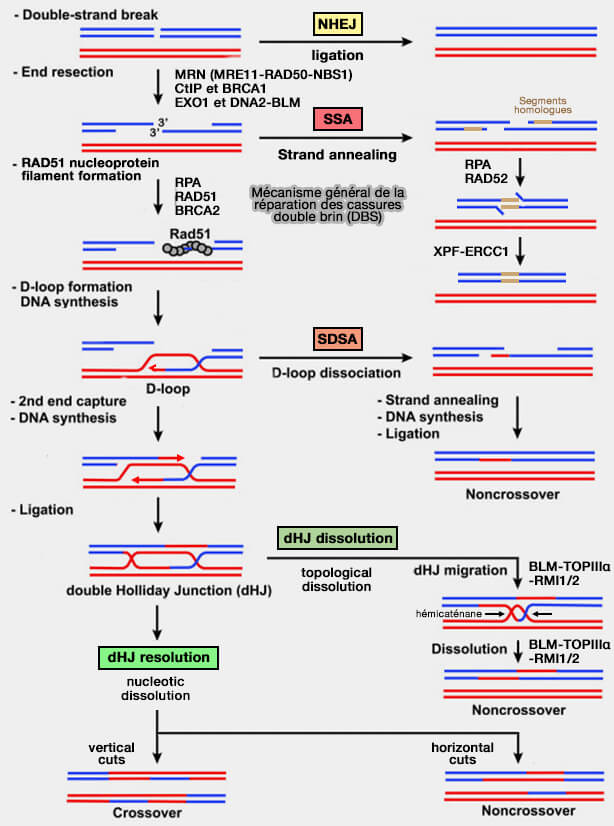
3. La réparation des cassures double brin repose sur plusieurs voies moléculaires distinctes qui diffèrent par leur mécanisme et leur fidélité et se répartissent en deux grandes stratégies.
a. Les mécanismes de recombinaison homologue utilisent une séquence d’ADN intacte comme matrice pour restaurer l’information génétique.
SSA (Single-Strand Annealing) est une voie de recombinaison dépendante d’homologie utilisant des séquences répétées situées de part et d’autre de la cassure, dont l’appariement conduit à la perte du segment intermédiaire.
b. Les mécanismes de jonction directe des extrémités d’ADN, qui religuent les extrémités cassées sans matrice homologue.
NHEJ (Non-Homologous End Joining) ligature directement les extrémités d’ADN et constitue une réparation rapide et active durant la majeure partie du cycle cellulaire, mais susceptible d’introduire des insertions ou délétions.
MMEJ/TMEJ (Microhomology-Mediated End Joining), voie alternative de jonction d’extrémités reposant sur de courtes micro-homologies et généralement associée à des délétions plus importantes.
Remarque : le terme alt-EJ (alternative end joining) désigne l’ensemble des mécanismes alternatifs de jonction des extrémités d’ADN indépendants du NHEJ classique dépendant de Ku, incluant notamment MMEJ/TMEJ et SSA, qui reposent sur l’utilisation de séquences homologues exposées après résection des extrémités.
HR est la seule voie réellement conservatrice, alors que NHEJ, MMEJ et SSA sont intrinsèquement mutagènes car ils entraînent souvent des insertions ou délétions.
4. Ces différentes voies constituent un réseau dynamique de réparation dont l’activation dépend :
l’absence de résection favorisant la jonction directe par NHEJ,
la résection 5'-3' des extrémités expose de l’ADN simple brin et oriente la réparation vers les mécanismes dépendant d’homologie tels que HR, SSA ou MMEJ.
Mécanisme global des cassures double brin de l’ADN (DSB, Double-Strand Breaks)
(Figure : vetopsy.fr d'après Daley et coll)
Recombinaison homologue (HR ou Homologous Recombination)
Principe
1. La recombinaison homologue (HR, Homologous Recombination) est un mécanisme de réparation des cassures double brin qui restaure fidèlement l’information génétique en utilisant une séquence d’ADN homologue intacte comme matrice (An Overview of the Molecular Mechanisms of Recombinational DNA Repair (2015).
Ce mécanisme repose le plus souvent sur la chromatide sœur produite lors de la réplication, ce qui permet de recopier la séquence correcte au niveau de la cassure.
La recombinaison homologue intervient principalement durant les phases S et G2 du cycle cellulaire, lorsque la chromatide sœur est disponible dans le noyau.
Remarque : cette contrainte explique que d’autres mécanismes de réparation des cassures double brin, notamment le NHEJ (Non-Homologous End Joining), soient privilégiés durant la phase G1, car aucune matrice homologue n’est disponible.
2. L’activation de la recombinaison homologue est également coordonnée par la réponse aux dommages de l’ADN, notamment par les kinases ATM et ATR, qui phosphorylent de nombreuses protéines impliquées dans la détection des cassures double brin, la résection des extrémités et l’assemblage des complexes de recombinaison, tout en régulant la progression du cycle cellulaire afin de permettre la réparation correcte des cassures double brin ( points de contrôle ou checkpoints).
L’activation des kinases ATM et ATR entraîne notamment la phosphorylation de la sérine 139 de la variante histonique H2AX, formant la marque γH2AX ( code des histones).
Cette modification chromatinienne s’étend sur de larges domaines autour de la cassure double brin et constitue une plateforme de recrutement pour de nombreuses protéines de la réponse aux dommages de l’ADN.
Mécanisme global
1. La cassure double brin génère deux extrémités d’ADN, chacune possédant un brin se terminant en 5' et un brin se terminant en 3', qui ne peuvent pas être réparées par simple recopie locale, car l’information génétique est perdue sur les deux brins au niveau de la rupture.
Remarque : la recombinaison peut également utiliser des séquences répétées homologues situées ailleurs dans le génome, phénomène, appelé recombinaison ectopique, qui peut provoquer des délétions, des duplications ou des réarrangements chromosomiques.
2. Dans le NHEJ (Non-Homologous End Joining), les extrémités d’ADN restent peu ou pas résectées, et peuvent être religuées directement.
Résection des extrémités de la cassure double brin
Contrôle de la résection et choix de la voie de réparation
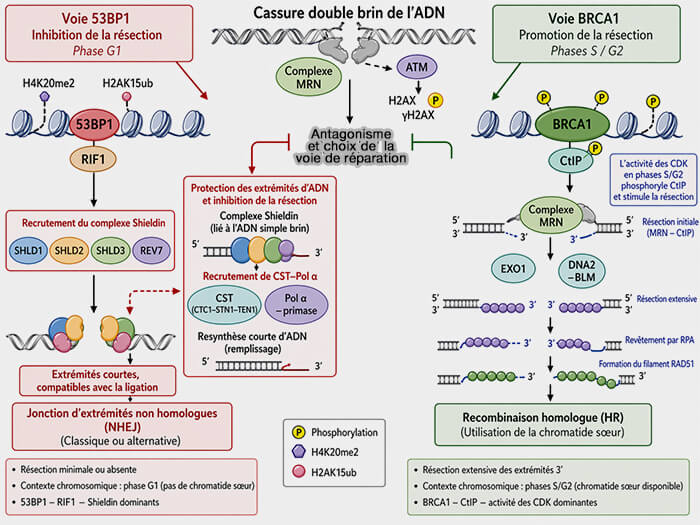
Le contrôle de la résection constitue l’un des principaux points de décision qui orientent la réparation des cassures double brin vers la recombinaison homologue (HR) ou vers la jonction d’extrémités non homologues (NHEJ).
Après la formation d’une cassure double brin, les extrémités d’ADN sont rapidement reconnues par le complexe MRN et par les facteurs de signalisation ATM qui phosphorylent l’histone H2AX (γH2AX) et déclenchent l’assemblage de plateformes de réparation chromatiniennes.
Une fois recrutée, 53BP1 agit comme un facteur anti-résection en recrutant notamment la protéine RIF1, qui à son tour favorise l’assemblage du complexe Shieldin composé des sous-unités SHLD1, SHLD2, SHLD3 et REV7.
Le complexe Shieldin possède une affinité pour l’ADN simple brin et agit en stabilisant les extrémités d’ADN générées au voisinage de la cassure.
En coopération avec le complexe CST (CTC1-STN1-TEN1) et la polymérase Pol α-primase, ce système peut même resynthétiser localement de courts segments d’ADN au niveau des extrémités résectées, ce qui limite l’extension de la résection et maintient les extrémités compatibles avec la ligation par la voie NHEJ.
NHEJ versus HR
(Figure : vetopsy.fr générée par IA)
2. Ce dispositif antagonise directement l’action de BRCA1 et l’équilibre fonctionnel entre les modules 53BP1-RIF1-Shieldin et BRCA1-CtIP constitue un mécanisme central de choix de la voie de réparation qui dépend également du contexte du cycle cellulaire.
Lorsque la résection est bloquée par 53BP1 et ses partenaires en l’absence de chromatide sœur en phase G1, les extrémités restent courtes et la réparation est dirigée vers la NHEJ.
L’activité des CDK en phase S/G2 favorise la phosphorylation de CtIP, ce qui stimule la résection initiale des extrémités d’ADN et facilite l’action de BRCA1, qui lève l’inhibition exercée par le module 53BP1-RIF1-Shieldin et permet l’engagement de la cellule dans la recombinaison homologue.
Remarque : la protéine 53BP1, décrite ici pour son rôle dans le contrôle de la résection des extrémités d’ADN et l’orientation de la réparation vers la voie NHEJ, peut également s’accumuler dans des corps nucléaires appelés 53BP1 nuclear bodies.
Ces domaines apparaissent principalement en phase G1 autour de régions chromatiniennes ayant subi un stress de réplication lors de la phase S précédente et participent à la surveillance de ces régions jusqu’à leur réparation.
Initiation de la résection des cassures double brin
1. Dans la recombinaison homologue, les extrémités d’ADN sont dégradées dans la direction 5' ➞ 3', ce qui génère des segments d’ADN simple brin possédant une extrémité 3'.
La résection des extrémités de la cassure constitue également un point de décision majeur dans le choix de la voie de réparation, et oriente la réparation vers la recombinaison homologue.
Remarque : dans la voie SSA (Single-Strand Annealing), deux séquences répétées homologues situées de part et d’autre de la cassure s’apparient directement après la résection des extrémités, ce qui entraîne la délétion de la région d’ADN comprise entre les répétitions.
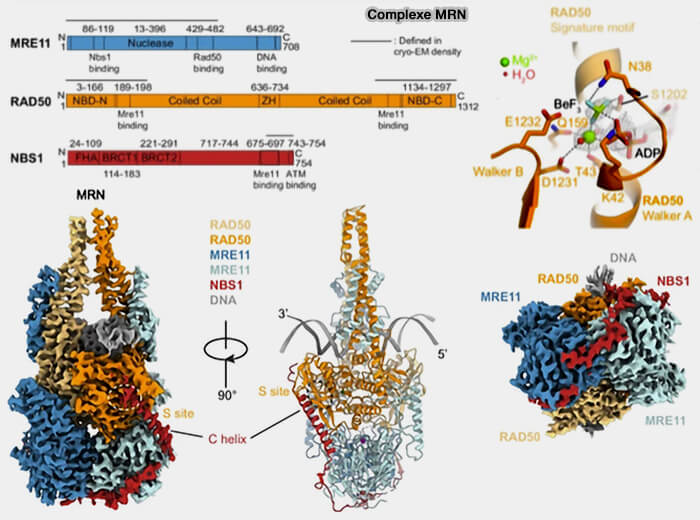
Structure du complexe MRN
(Figure : vetopsy.fr d'après Fan et coll)
la protéine MRE11 qui possède une activité nucléase et participe au traitement initial des extrémités d’ADN,
RAD50 qui assure l’architecture du complexe et maintient les extrémités cassées à proximité grâce à ses domaines coiled-coil et à son domaine en anneau qui permet le rapprochement des deux extrémités de la cassure,
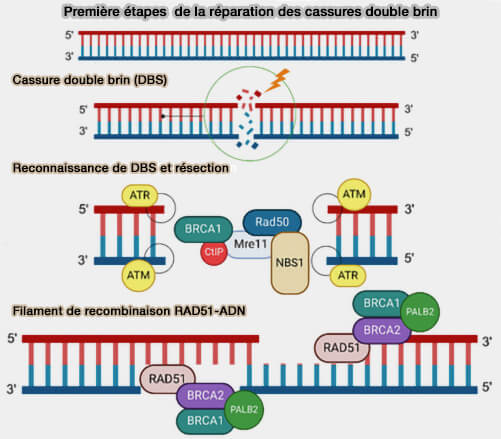
Premières étapes de la réparation des cassures double brin
(Figure : vetopsy.fr d'après Panagopoulou et coll)
3. BRCA1 (BReast CAncer 1) favorise également la résection des extrémités en facilitant l’activité de la protéine CtIP (CtBP-interacting protein), protéine en grande partie intrinsèquement désordonnée qui interagit avec le domaine BRCT (BRCA1 C-Terminal) de BRCA1 et stimule l’activité nucléase du complexe MRN au voisinage de la cassure.
a. Le domaine BRCT (BRCA1 C-Terminal) est un domaine d’interaction protéique présent dans plusieurs protéines impliquées dans la réponse aux dommages de l’ADN (DDR).
Il reconnaît généralement des motifs phosphorylés et on le retrouve notamment dans BRCA1, LIG3, XRCC1 et CtIP, où il contribue au recrutement et à la stabilisation des complexes impliqués dans la réparation des cassures de l’ADN.
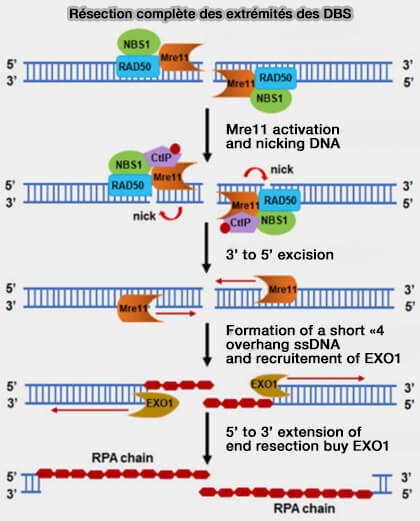
Résection complète des extrémités des DBS
(Figure : vetopsy.fr d'après Yan et coll)
Bien que BRCA1 soit principalement connue pour son rôle dans la réparation des cassures double brin et la stabilité du génome, ses domaines BRCT peuvent également interagir avec certaines enzymes métaboliques comme ACC1 (Acétyl-CoA carboxylase), reliant ainsi la régulation du cycle cellulaire et le métabolisme lipidique.
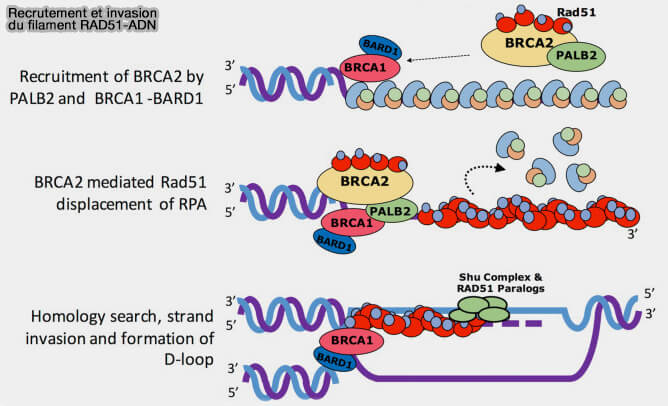
b. BRCA1 forme un hétérodimère stable avec BARD1 via leurs domaines RING, et ce complexe BRCA1-BARD1 participe au recrutement des facteurs de recombinaison homologue, notamment PALB2 et BRCA2, facilitant ainsi le chargement de RAD51 sur l’ADN simple brin.
a. CtIP, dont l’activité est régulée par phosphorylation par les kinases du cycle cellulaire (CDK) et de la réponse aux dommages de l’ADN (ATM), constitue ainsi un facteur clé favorisant l’engagement de la réparation vers la recombinaison homologue.
b. Cette activation conduit à une incision du brin 5′ par MRE11, mais qui n’est pas réalisée exactement au niveau de la cassure, mais à une certaine distance sur le brin 5′.
À partir de ce point d’incision, MRE11 utilise son activité exonucléase 3′ → 5′ pour dégrader l’ADN en direction de la cassure, ce qui permet le traitement initial des extrémités.
Résection étendue et stabilisation de l’ADN simple brin
1. La résection est ensuite étendue sur de longues distances par des nucléases et des hélicases spécialisées selon deux voies principales.
Dans le système DNA2-BLM, l’hélicase BLM déroule l’ADN tandis que la nucléase DNA2 dégrade le brin déplacé.
La résection génère finalement de longues queues d’ADN simple brin possédant une extrémité 3' nécessaire à la recombinaison homologue.
2. Les régions d’ADN simple brin générées par la résection sont immédiatement recouvertes par RPA (Replication Protein A).
Elle se fixe rapidement sur les queues 3' simple brin produites par la résection et stabilise ces régions d’ADN afin d’empêcher la formation de structures secondaires ou leur dégradation (stabilisation de l'ADN simple brin par RPA).
RPA empêche l’appariement non spécifique des séquences et maintient l’ADN dans une conformation accessible aux protéines de recombinaison.
Remarque : ATR est activée après la résection, lorsque l’ADN simple brin est recouvert par RPA, pour stabiliser la réparation par recombinaison homologue et coordonner la réparation avec le cycle cellulaire, en phosphorylant notamment Chk1, BRCA1, PALB2, plusieurs facteurs du cycle cellulaire, Cdc25A, CDC25C et Wee1 pour l'inhibition CDC25/activation WEE1 qui bloque le cycle cellulaire pendant la réparation de l’ADN (checkpoint G2/M).
La recombinaison homologue nécessite ensuite le remplacement de RPA par la recombinase RAD51 (RAD51: Beyond the break 2021).
1. La structure monomérique de Rad51 est constituée par :
un domaine N-terminal impliqué dans les interactions entre protomères de RAD51 lors de la polymérisation du filament, contribuant à la stabilité du filament nucléoprotéique,
a. Chaque protomère interagissant avec l’ADN simple brin par l’intermédiaire de boucles de liaison à l’ADN (L1 et L2) qui stabilisent le filament.
b. L’ADN simple brin est :
étiré et rigidifié (~1,5 de sa longueur normale),
organisé en triplets de bases exposés vers l’extérieur, configuration qui permet de comparer simultanément trois bases du brin simple brin avec trois bases du duplex d’ADN testé.
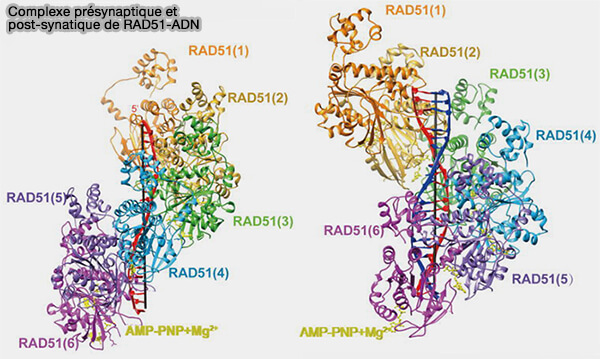
Complexe présynaptique et postsynaptique
(Figure : vetopsy.fr d'après Fan et coll)
3. Dans la recombinaison homologue, on distingue trois états structuraux successifs du filament de recombinaison RAD51-ADN.
Le complexe présynaptique correspond au filament nucléoprotéique RAD51-ADN simple brin formé après la résection des extrémités qui réalise la recherche d’homologie en sondant les molécules d’ADN double brin voisines.
Le complexe synaptique correspond à l’état transitoire dans lequel le filament RAD51-ADN établit un contact avec une molécule d’ADN double brin et teste l’appariement des bases entre l’ADN simple brin du filament et le duplex.
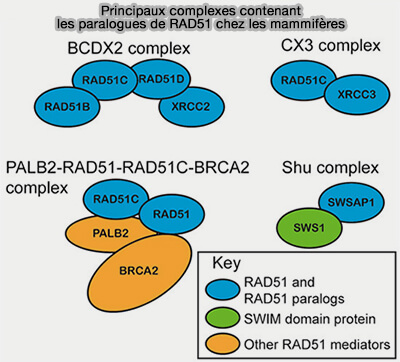
Paralogues de RAD51
(Figure : vetopsy.fr d'après Sullivan et coll)Le complexe postsynaptique correspond à l’état où le simple brin envahit le duplex homologue et s’apparie avec le brin complémentaire, ce qui conduit à la formation d’une D-loop et initie la synthèse d’ADN réparatrice.
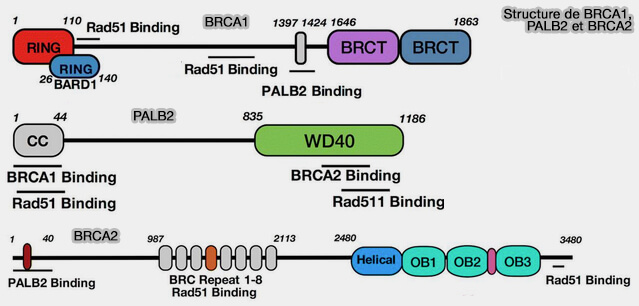
Cette activité est facilitée par la protéine PALB2 (Partner And Localizer of BRCA2), qui interagit directement avec le domaine coiled-coil de BRCA1 et avec BRCA2, permettant le recrutement de BRCA2 aux sites de cassure et stabilisant le complexe de chargement de RAD51.
BRCA2 interagit directement avec RAD51 via ses motifs BRC, BRCA2 charge RAD51 sur l’ADN simple brin pour former le filament RAD51-ADN, essentiel pour la recombinaison homologue.
b. Les paralogues de RAD51 (RAD51B, RAD51C, RAD51D, XRCC2 et XRCC3), qui forment généralement deux complexes, BCDX2 et CX3, stabilisent le filament RAD51 et facilitent son assemblage sur l’ADN simple brin (RAD51 paralog function in replicative DNA damage and tolerance 2022).
Structures de BRCA1, PALB2 et BRCA2
(Figure : vetopsy.fr d'après Sun et coll)
Invasion de brin, formation de la D-loop et synthèse d’ADN
Le filament RAD51-ADN simple brin recherche l’homologie dans la chromatide sœur en interagit transitoirement avec des molécules d’ADN double brin voisines pour tester l’appariement des bases.
1. Cette recherche d’homologie repose sur la structure particulière du filament RAD51-ADN.
a. L’ADN simple brin est étiré et organisé en triplets de bases exposés vers l’extérieur, tandis que les boucles L1 et L2 des protomères RAD51 interagissent avec l’ADN.
Le filament peut ainsi établir des contacts transitoires avec le sillon majeur du duplex, ce qui induit une déformation et un étirement local de l’ADN double brin.
Le duplex est transitoirement ouvert et les bases du simple brin du filament peuvent comparer leur complémentarité avec celles du duplex.
b. Ces interactions permettent au filament de tester l’homologie par segments courts (~8–15 nucléotides).
2. Deux situations peuvent alors se produire.
Si l’appariement est incorrect, l’interaction est instable et le duplex d’ADN se referme spontanément en retrouvant sa conformation thermodynamiquement stable.
Lorsque l’homologie est suffisante, l’appariement est stabilisé et le brin simple envahisseur s’insère dans le duplex homologue, ce qui initie l’invasion de brin et la formation de la D-loop.
Recrutement et invasion du filament RAD51-ADN
(Figure : vetopsy.fr d'après Sun et coll)
a. Le brin simple 3' envahisseur, maintenu dans le filament RAD51, est présenté au duplex homologue dans une conformation étirée et préorganisée.
Les interactions du filament avec l’ADN double brin provoquent un étirement et une déstabilisation locale du duplex, ce qui facilite la séparation transitoire des deux brins.
Dans ces conditions, les bases du simple brin peuvent former des appariements Watson-Crick avec le brin complémentaire du duplex, tandis que les boucles L1 et L2 de RAD51 stabilisent ce nouvel appariement.
b. Le brin envahisseur remplace progressivement le brin initialement apparié du duplex, qui est repoussé vers l’extérieur.
Cette interaction provoque le déplacement local de l’autre brin du duplex, qui reste attaché à la molécule d’ADN mais forme une boucle d’ADN simple brin.
c. L’ensemble forme une structure intermédiaire appelée D-loop (displacement loop), dans laquelle le brin envahisseur est apparié à la matrice homologue tandis que le brin déplacé constitue la boucle simple brin caractéristique.
Remarque : une structure de type D-loop est également observée aux télomères, où l’extrémité 3' simple brin du télomère (organisation et structure des télomères)
4. Une ADN polymérase, principalement la polymérase δ et parfois la polymérase ε, allonge ensuite l’extrémité 3' envahissante en copiant la séquence de la chromatide sœur, ce qui permet de restaurer progressivement l’ADN manquant au niveau de la cassure.




 recombinaison méiotique),
recombinaison méiotique),