L’élongation de la réplication repose sur la synthèse continue du brin directeur et la synthèse discontinue du brin retardé, suivie de la maturation des fragments d’Okazaki.
Ces étapes correspondent aux différents mécanismes moléculaires qui assurent la progression et l’achèvement de la réplication du génome (terminaison de la duplication).
Cette organisation positionne la polymérase ε immédiatement derrière l’hélicase, ce qui permet la synthèse continue du brin directeur à mesure que la double hélice est déroulée.
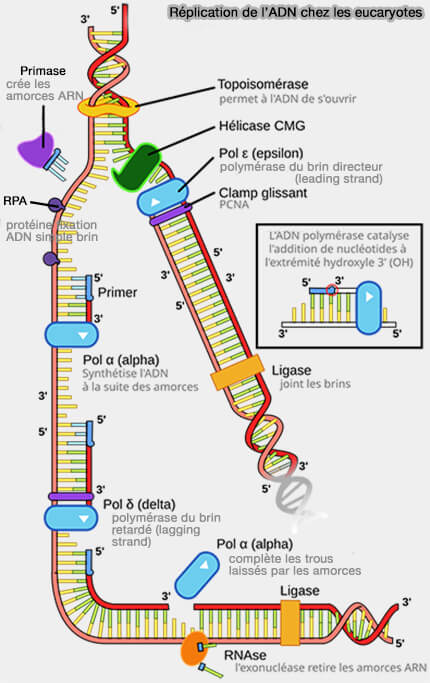
Synthèse dans le brin directeur et retardé de la fourche de réplication eucaryote
(Figure : vetopsy.fr d'après Nasheuer et coll)
1. La polymérase reste associée à l’ADN grâce au clamp de processivité PCNA (Proliferating Cell Nuclear Antigen), l'anneau protéique trimérique qui entoure l’ADN et stabilise l’enzyme sur la matrice pendant l’élongation.
PCNA agit comme un support coulissant permettant à la polymérase de rester attachée à l’ADN tout en progressant le long de la matrice, ce qui augmente considérablement la processivité de la synthèse.
2. La protéine Mrc1 (Mediator of Replication Checkpoint 1), appelée Claspin chez les vertébrés, est associée au réplisome et interagit à la fois avec l’hélicase CMG et avec l’ADN polymérase ε du brin directeur.
a. Elle contribue ainsi à coupler l’ouverture de la double hélice par l’hélicase avec la synthèse continue du brin directeur, favorisant une progression coordonnée de la fourche de réplication.
Lorsque Mrc1 est absent, la fourche progresse beaucoup plus lentement et la stabilité du réplisome est altérée.
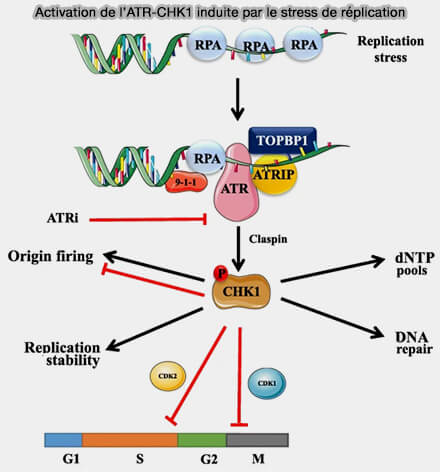
Activation de l’ATR-CHK1 induite par le stress de réplication
(Figure : vetopsy.fr d'après Mei et coll)
b. Mrc1 participe également à la stabilisation de la fourche et joue un rôle important dans la réponse aux perturbations de la réplication en transmettant les signaux de stress réplicatif aux mécanismes de surveillance du cycle cellulaire au niveau du checkpoint S.
Lorsque la fourche rencontre un problème, par exemple un manque de nucléotides, des lésions de l’ADN ou un blocage de la synthèse, Mrc1 est phosphorylée et transmet le signal de stress aux kinases de checkpoint, notamment Rad53 chez la levure/Chk1 chez les vertébrés.
L’activation de ces voies de signalisation permet de ralentir la progression de la réplication, de stabiliser la fourche et de laisser le temps aux mécanismes de réparation de l’ADN d’agir.
Cette organisation permet à la polymérase δ de se dissocier et de se réassocier successivement à l’ADN au cours de la synthèse des différents fragments d’Okazaki.
Synthèse des fragments d’Okazaki
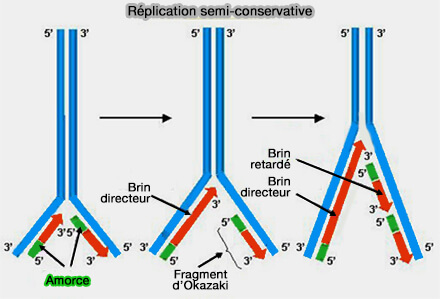
Synthèse discontinue du brin retardé
La synthèse discontinue du brin retardé conduit à la formation de segments courts d’ADN appelés fragments d’Okazaki.
Réplication de l'ADN
(Figure : vetopsy.fr d'après LadyofHats)
1. Chez les eucaryotes, les fragments d’Okazaki mesurent généralement environ 100 à 200 nucléotides, une longueur proche de celle de l’ADN associé à un nucléosome.
Lors de la progression de la fourche de réplication, les nucléosomes situés en aval constituent des obstacles transitoires pour la polymérase δ.
La synthèse d’un fragment d’Okazaki se poursuit jusqu’à proximité du nucléosome suivant, après quoi une nouvelle amorce est synthétisée en amont.
Cette organisation reflète ainsi l’influence directe de la structure nucléosomique de la chromatine sur la longueur des fragments d’Okazaki et sur la progression de la synthèse du brin retardé.
2. Chaque fragment initié par l'amorce ARN-ADN est allongé par l’ADN polymérase δ.
Chaque nouveau fragment nécessite également le chargement d’un nouveau clamp PCNA, qui assure la processivité de la polymérase δ au cours de l’élongation.
3. La synthèse discontinue du brin retardé et le fait que la polymérase δ doit synthétiser l’ADN dans le sens 5' ➞ 3', donc dans une direction opposée à la progression de la fourche de réplication, imposent une organisation particulière de la fourche de réplication.
a. Dans le réplisome, les deux polymérases doivent être maintenues près de l’hélicase CMG qui sépare les deux brins parentaux, avance le long de l’ADN et génère en permanence de nouvelles régions simple brin.
Si la polymérase du brin retardé synthétisait simplement l’ADN, elle partirait loin derrière la fourche, se séparerait du réplisome et ne pourrait plus recevoir la prochaine amorce.
b. Pour rester associée au réplisome tout en synthétisant les fragments d’Okazaki, le brin matrice retardé forme une boucle transitoire qui replie l’ADN vers le complexe réplicatif.
Cette boucle permet à la polymérase δ d’allonger un fragment d’Okazaki tout en restant orientée dans la même direction globale que la polymérase ε du brin directeur.
L'organisation coordonnée du réplisome permet d’optimiser la progression de la fourche de réplication et d’assurer la synchronisation de la synthèse des deux brins d’ADN.
Mécanisme de la synthèse
Le mécanisme simplifié est le suivant.
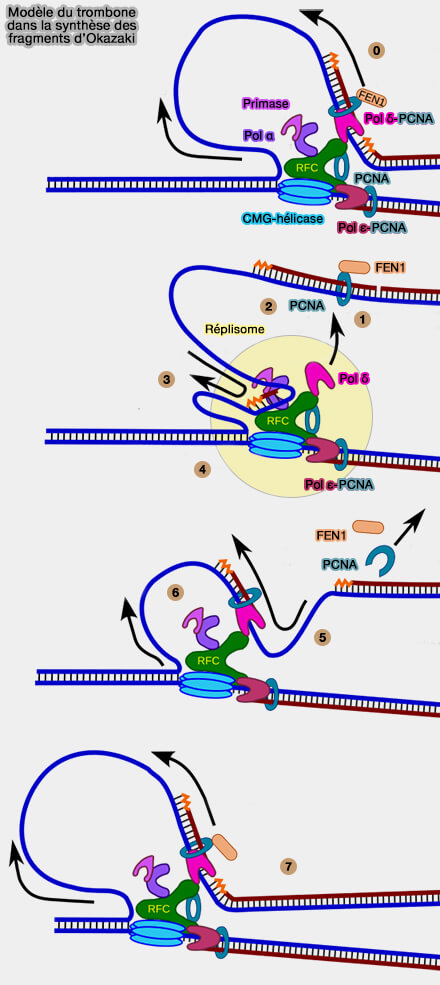
Modèle du trombone dans la synthèse des fragments d’Okazaki
(Figure : vetopsy.fr d'après RWhitwam)
0. Le fragment d’Okazaki en cours d’élongation est presque terminé.
Le brin matrice retardé est tiré à travers le complexe Pol δ-PCNA, ce qui permet à la polymérase δ de continuer la synthèse du nouveau fragment.
Pendant ce temps, l’hélicase CMG continue d’ouvrir l’ADN et un nouveau segment de matrice simple brin apparaît.
1. Lorsque la polymérase δ atteint le fragment précédent, la synthèse du fragment d’Okazaki se termine.
La polymérase δ se dissocie alors du fragment nouvellement synthétisé.
Le clamp PCNA reste associé à l’ADN et participe au recrutement des enzymes impliquées dans la maturation du fragment, notamment FEN1.
2. Le primosome (primase + Pol α) capture le brin matrice retardé près de la fourche et initie la synthèse d’un nouveau fragment.
Il forme une boucle d’ADN d'environ 150 à 200 nucléotides en amont du fragment terminé, qui correspond approximativement à la longueur typique des fragments d’Okazaki chez les eucaryotes.
3. Le brin matrice retardé est tiré à travers la primase qui catalyse la synthèse d’une courte amorce ARN.
4. Le brin matrice retardé est transmis à l’ADN polymérase α qui synthétise une courte extension d’ADN, formant l’amorce ARN-ADN qui servira de point de départ pour le nouveau fragment d’Okazaki.
5. Le primosome se détache du brin retardé.
Le facteur de chargement RFC recrute ensuite PCNA et l’ouvre pour le placer autour de l’ADN.
PCNA forme alors le clamp de processivité du nouveau fragment.
6. Le complexe PCNA-ADN est transféré à la polymérase δ. Pol δ se fixe sur PCNA et devient la polymérase responsable de l’élongation du nouveau fragment d’Okazaki.
7. La polymérase δ recommence l’élongation.
Le brin matrice retardé est tiré à travers Pol δ-PCNA, ce qui entraîne la croissance de la boucle du trombone.
La synthèse se poursuit jusqu’à ce que la polymérase atteigne le fragment précédent, ce qui ramène le système à l’étape 0 du cycle suivant.
Remarque : ce schéma illustre le modèle pédagogique du trombone, dans lequel les polymérases du brin directeur et du brin retardé apparaissent associées de manière stable au réplisome.
Chez les eucaryotes, les données structurales indiquent cependant que la polymérase δ est recrutée de façon dynamique pour chaque fragment d’Okazaki via PCNA, plutôt que d’être fixée en permanence au complexe réplicatif.
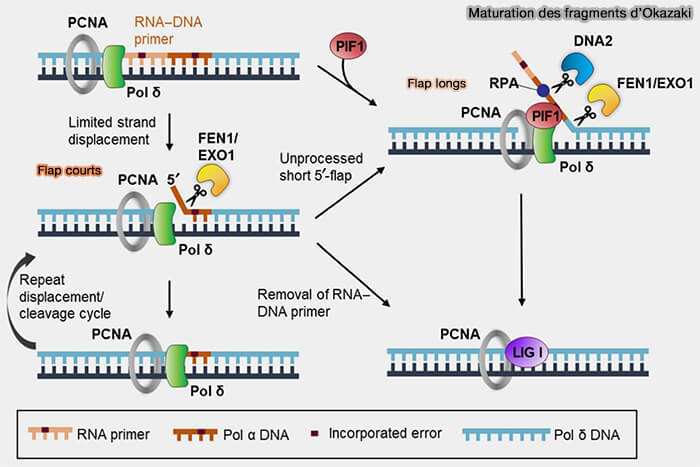
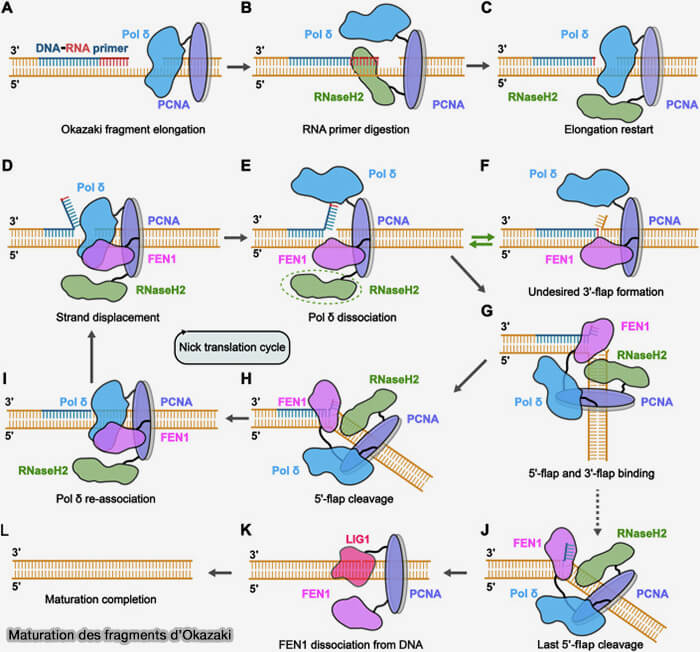
1. La maturation débute lorsque la polymérase δ synthétisant le fragment d’Okazaki en cours atteint l’amorce ARN du fragment précédent.
Lors de la synthèse du fragment d’Okazaki suivant, la polymérase δ poursuit brièvement l’élongation et déplace l’extrémité 5′ du fragment précédent (activité de déplacement de brin ou strand displacement), ce qui refoule le segment contenant l’amorce et génère un segment d’ADN simple brin appelé flap 5′.
Maturation des fragments d'Okazaki
(Figure : vetopsy.fr d'après Sun et coll)
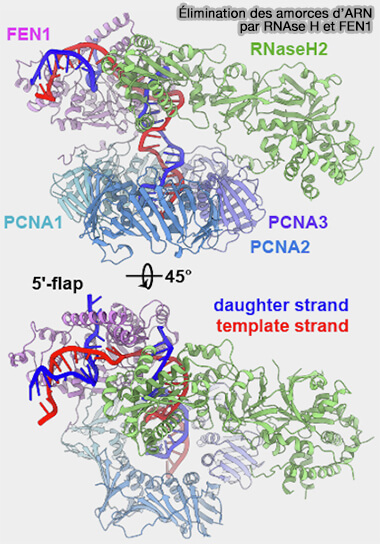
Élimination des amorces d’ARN par RNAse-H et FEN1
(Figure : vetopsy.fr d'après Tian et coll)
a. La RNase H hydrolyse la majeure partie des ribonucléotides constituant l’amorce d’ARN hybridée à l’ADN, mais laisse généralement subsister le dernier ribonucléotide situé à la jonction ARN-ADN. (Ribonuclease H: the enzymes in Eukaryotes 2009).
La RNase H n’est pas représentée sur ce schéma car celui-ci se concentre essentiellement sur les mécanismes de traitement des flaps générés par le déplacement de brin lors de la synthèse par la polymérase δ.
b. Ce flap contenant les nucléotides résiduels de l’amorce est reconnu et clivé par l’endonucléase FEN1 (Flap Endonuclease 1), qui élimine le segment déplacé et rétablit une extrémité d’ADN compatible avec la jonction des fragments (Flap Endonuclease 1 2013).
Ces flaps longs peuvent être favorisés par l’action de PIF1, hélicase 5' → 3', qui peut dérouler l’ADN déplacé par la polymérase δ, ce qui favorise la formation de flaps plus longs (Structure and Function of Pif1 Helicase 2017).
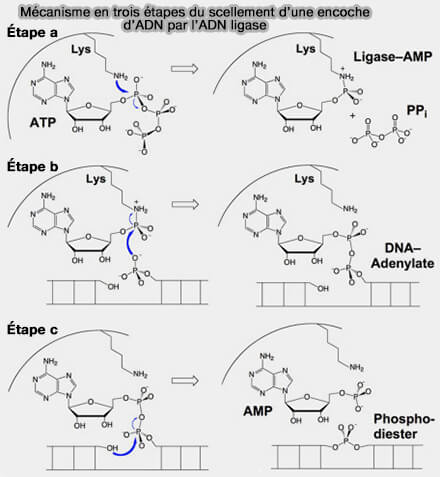
Dans le site actif de LIG1, un résidu lysine catalytique conservé (Lys568 chez l’humain) attaque le phosphate α de l’ATP.
Il se produit la formation d’un intermédiaire covalent ligase-AMP par liaison phosphoramidate entre l’AMP et la lysine catalytique, accompagnée de la libération de pyrophosphate (PPi), l’AMP servant ensuite de groupe activateur du substrat ADN.
b. Lorsque LIG1 reconnaît une encoche pouvant être scellée (3′-OH et 5′-phosphate correctement appariés), l’enzyme se referme autour de l’ADN et l’AMP lié à la lysine est transféré au phosphate 5′ du brin aval, ce qui génère un intermédiaire appelé ADN-adénylate (AppDNA).
c. Le phosphate 5′ activé devient alors un phosphate adénylé beaucoup plus réactif qui est attaqué par le groupement 3′-OH du brin amont, ce qui entraîne la formation de la liaison phosphodiester 3′-5′ et la libération de l’AMP.
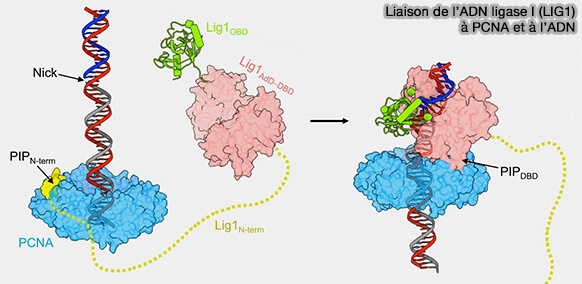
Liaison de l’ADN ligase I (LIG1) à PCNA et à l’ADN
(Figure : vetopsy.fr d'après Blair et coll)
a. La ligase reste liée au clamp PCNA tout en conservant une certaine mobilité, ce qui lui permet d’explorer l’ADN adjacent afin de localiser une jonction simple brin (entaille ou nick) générée après l’action de Pol δ et de FEN1 lors de la maturation des fragments d’Okazaki.
Lorsque LIG1 rencontre une entaille comportant un 3′-OH et un 5′-phosphate, elle se fixe directement sur l’ADN et l’enzyme se referme autour du duplex.
La structure montre alors un assemblage dans lequel PCNA forme un premier anneau entourant l’ADN tandis que la ligase entoure également partiellement l’ADN au niveau de l'enrtaille, créant une architecture stabilisée par des interactions supplémentaires entre PCNA et le domaine de liaison à l’ADN de LIG1.
b. Dans ce modèle dit “ toolbelt ”, FEN1 et LIG1 peuvent donc être associées en même temps à différentes sous-unités de PCNA.
FEN1, attachée à une première sous-unité de PCNA via son motif PIP C-terminal, est positionnée de manière à accéder au flap 5′ généré par l’activité de déplacement de brin de la polymérase δ.
La DNA ligase I (LIG1) est recrutée sur une deuxième sous-unité de PCNA, via son motif PIP N-terminal, ce qui maintient la ligase à proximité du site où FEN1 vient de cliver le flap.
La troisième sous-unité de PCNA peut rester libre ou être occupée par une autre protéine du réplisome, comme Pol δ, qui reste associée au clamp pendant la synthèse et le traitement du fragment d’Okazaki.
c. Cette réorganisation de l’interaction avec PCNA permet d’aligner précisément le site catalytique de la ligase sur la jonction à sceller, condition nécessaire à la formation de la liaison phosphodiester.
La réplication de l’ADN s’achève lorsque les fourches de réplication convergent et que les derniers segments d’ADN sont synthétisés puis maturés, assurant la continuité des molécules filles.
La fidélité de cette synthèse est contrôlée par les mécanismes de correction d’épreuve (proofreading) des polymérases réplicatives, qui éliminent les nucléotides incorrectement incorporés afin de maintenir l’intégrité du génome.




{kind=link}
 terminaison de la duplication).
terminaison de la duplication). 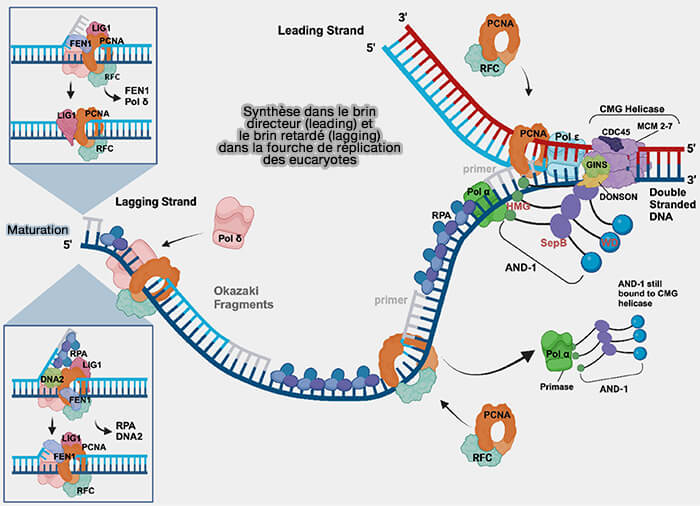
{kind=link}

{kind=link}