


Acides nucléiques
ADN : réparation
sans cassure double brin
Réparation des mésappariements (MMR)
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Le système de réparation des mésappariements (MMR) corrige les erreurs d’appariement et les insertions ou délétions apparues lors de la réplication de l’ADN afin de maintenir la fidélité du génome.
La réparation de l’ADN correspond spécifiquement à la correction enzymatique des lésions affectant la molécule d’ADN et repose sur plusieurs systèmes spécialisés :
a. réparant des lésions de l’ADN sans cassure double brin :
b. impliquant des cassures double brin (DSBR) :
- HR (recombinaison homologue)
- la réparation par jonction des extrémités d’ADN (NHEJ, SSA, MMEJ/TMEJ).
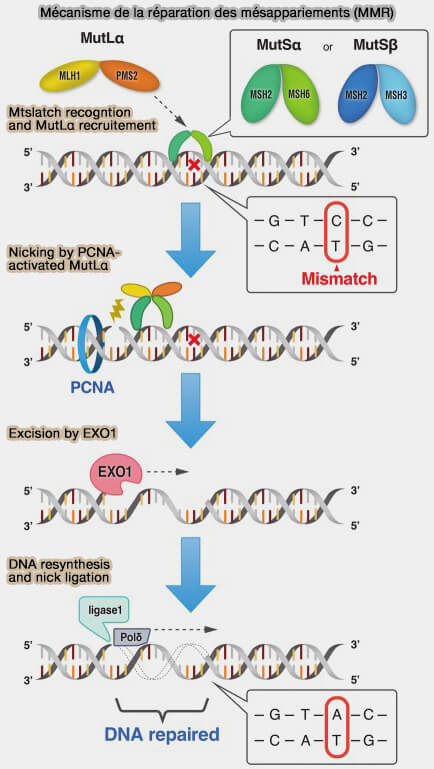
Principe de la réparation des mésappariements (MMR)
1. La réparation des mésappariements (MMR ou Mismatch Repair) constitue l’un des principaux mécanismes de fidélisation de la réplication de l’ADN.
a. Malgré la haute fidélité des ADN polymérases et l’activité de relecture 3' → 5' (proofreading), certaines erreurs persistent, notamment des mésappariements de bases (A-C, G-T…) ainsi que des insertions ou délétions d’un ou plusieurs nucléotides générées lors du glissement de la polymérase sur les séquences répétées.
(Figure : vetopsy.fr d'après biorender.com)
Les boucles d’insertion ou de délétion générées lors de ce glissement constituent une source majeure d’erreurs dans les séquences répétées du génome, notamment les microsatellites.
b. En corrigeant les erreurs d’appariement résiduelles introduites par ces enzymes lors de la réplication, le système MMR réduit fortement le taux global de mutation et contribue au maintien de la stabilité du génome.
2. Le système MMR repose sur la détection des distorsions locales de la géométrie de la double hélice d’ADN provoquées par un mauvais appariement de bases.
Ces altérations structurales constituent le signal primaire permettant au système de réparation d’identifier la présence d’une erreur dans le duplex.
3. Une fois l’erreur détectée, la réparation doit cibler le brin néosynthétisé, tandis que le brin parental intact sert de matrice pour restaurer la séquence correcte.
Chez les eucaryotes, le mécanisme repose principalement sur des signaux associés à la réplication, notamment les discontinuités transitoires du brin néosynthétisé telles que :
- les fragments d’Okazaki du brin retardé,
- les discontinuités transitoires générées lors de la synthèse du brin avancé.
Remarque : chez les bactéries, la discrimination entre les deux brins repose sur un mécanisme différent fondé sur l’état de méthylation de l’ADN. Dans le système MutHLS dépendant de la méthylation Dam, la réparation est dirigée vers le brin néosynthétisé qui n’est pas encore méthylé.
4. Après la reconnaissance d’un mésappariement et l’identification du brin néosynthétisé, la réparation est assurée par une série de complexes protéiques conservés au cours de l’évolution, notamment MutS, MutL et la pince de réplication PCNA qui contribue à orienter la réparation vers le brin récemment synthétisé.
Reconnaissance du mésappariement
1. Chez les eucaryotes, deux hétérodimères principaux interviennent dans la reconnaissance du mésappariement.
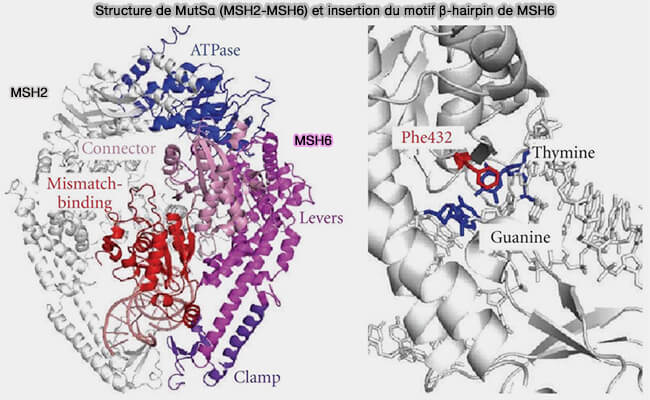
a. MutSα (MSH2-MSH6) et MutSβ (MSH2-MSH3) sont des complexes apparentés aux protéines bactériennes MutS, qui reconnaissent principalement la distorsion structurale du duplex plutôt que la nature chimique des bases.
- MutSα reconnaît principalement les mésappariements simples comme G-T ou A-C et les petites boucles d’insertion/délétion (IDL) d'un nucléotide.
- MutSβ reconnaît préférentiellement les boucles d’insertion/délétion plus longues générées lors du glissement de la polymérase sur les séquences répétées.
(Figure : vetopsy.fr d'après Zhang et coll)
b. La reconnaissance du mésappariement implique l’insertion d’un motif β-hairpin provenant de la sous-unité MSH6 pour MutSα ou MSH3 et la sous-unité MSH3 pour MutSβ dans le sillon mineur de l’ADN.
- Cette insertion stabilise une déformation locale du duplex, caractérisée par un basculement des bases mal appariées et un élargissement local du sillon mineur lié à la courbure de l’ADN et à l’extrusion partielle de la base incorrectement appariée.
- Cette interaction agit comme un capteur structural permettant au complexe MutS de distinguer un mésappariement d’un appariement canonique.
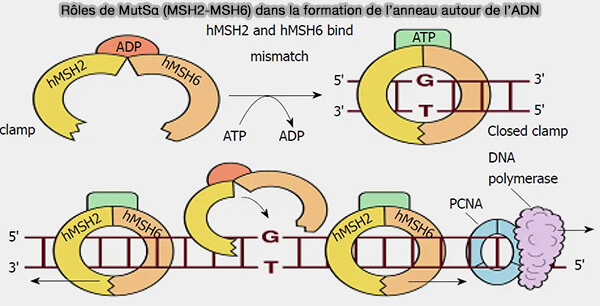
2. Après reconnaissance du site erroné, MutSα subit une transition conformationnelle dépendante de l’ATP.
a. La fixation d’ATP dans les domaines ATPase des protéines MSH induit une transition conformationnelle du complexe MutS, la β-hairpin se retire du sillon mineur et le complexe MutS adopte une conformation fermée en anneau autour de l’ADN pour devenir un clamp topologique.
Cette configuration, dite " sliding clamp " capable de glisser le long de l’ADN, est due à l'absence d'interaction forte avec l’ADN, les contacts devenant principalement électrostatiques et non spécifiques.
b. Ce processus, appelé diffusion unidimensionnelle le long de l’ADN, résulte des fluctuations thermiques qui déplacent l’ADN dans le canal central du clamp MutS (≈25 Å), légèrement plus large que la double hélice (≈20 Å), permettant la diffusion du complexe tout en le maintenant topologiquement associé à l’ADN.
- À l’échelle moléculaire, toutes les molécules subissent des agitations thermiques permanentes dues aux collisions avec les molécules du solvant qui, ici, poussent légèrement le complexe et déplacent l’ADN dans le canal du clamp.
- La diffusion est dite unidimensionnelle parce que le complexe ne peut pas quitter l’ADN (anneau) et ne peut se déplacer que le long de l’axe de la double hélice.
(Figure : vetopsy.fr d'après Zhang et coll)
3. Ce changement de conformation libère partiellement le site du mésappariement tout en maintenant le complexe MutS associé à l’ADN.
- Le site d’erreur peut alors être reconnu à nouveau par un second complexe MutS, qui se fixe à son tour sur le mésappariement.
- Ainsi, plusieurs complexes MutS peuvent être chargés successivement au niveau du site d’erreur.
4. Cette accumulation locale de clamps favorise le recrutement efficace du complexe MutL, qui est ensuite chargé sur l’ADN et déclenche l’étape d’incision du brin néosynthétisé.
Activation de la réparation par MutLα et couplage avec PCNA
Chez les eucaryotes, la réparation des mésappariements est étroitement couplée à la réplication de l’ADN.
1. Ce couplage repose sur l’interaction du système MMR avec le clamp de réplication PCNA (Proliferating Cell Nuclear Antigen) qui entoure l’ADN et assure la processivité des ADN polymérases.
- PCNA constitue un anneau trimérique chargé sur l’ADN par le facteur RFC lors de la réplication.
- Sa structure impose une polarité fixe sur la double hélice, ce qui permet au système MMR de distinguer le brin néosynthétisé du brin matrice et d’orienter la réparation vers ce brin.

Vous pouvez lire : MutS functions as a clamp loader by positioning MutL on the DNA during mismatch repair (2022).
2. Lorsqu’un mésappariement est détecté, le complexe MutSα se fixe sur la distorsion locale de l’ADN et recrute le complexe MutLα (MLH1-PMS2), qui constitue l’effecteur central de la réparation (Endonuclease activities of MutLα and its homologs in DNA mismatch repair 2017).
La sous-unité PMS2 possède une activité endonucléase qui incise le brin néosynthétisé, initiant l’excision du segment d’ADN contenant l’erreur avant sa resynthèse.
3. Le complexe MutLα agit ensuite comme une plateforme d’intégration des signaux de réparation et interagit notamment avec la pince de réplication PCNA, contribuant à orienter la réparation vers le brin récemment synthétisé (MutS/MutL crystal structure reveals that the MutS sliding clamp loads MutL onto DNA 2015).
- Cette interaction déclenche une transition conformationnelle du complexe MutLα dans laquelle les domaines N-terminaux ATPasiques des sous-unités MLH1 et PMS2 se rapprochent et se referment autour de l’ADN pour former un anneau protéique topologiquement associé à la double hélice.
- Ce mécanisme est analogue à celui observé pour MutS : la fixation d’ATP stabilise l’association des deux sous-unités et provoque la fermeture du clamp MutL autour de l’ADN.
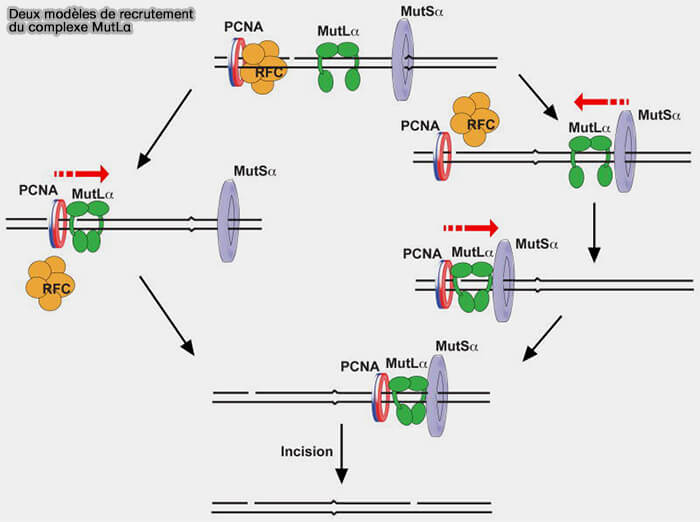
(Figure : vetopsy.fr d'après Kadyrova et coll)
4. Dans cette configuration, l’interaction avec PCNA active l’activité endonucléase portée par la sous-unité PMS2.
En raison de la polarité structurale de PCNA sur l’ADN, le complexe MutLα est positionné asymétriquement par rapport au mismatch, ce qui permet à l’endonucléase PMS2 d’introduire une coupure dans le brin néosynthétisé, à proximité du mésappariement.
5. Cette incision constitue le signal initial de l’étape d’excision, fournissant le point d’entrée pour les enzymes qui élimineront le segment du brin néosynthétisé contenant l’erreur avant la resynthèse correcte de l’ADN.
Excision du segment contenant l’erreur et resynthèse de l’ADN
1. L’excision du segment d’ADN contenant le mésappariement implique le déroulement local du duplex et la dégradation directionnelle du brin cible jusqu’au site d’erreur par l’exonucléase EXO1 recrutée par le complexe MutLα, ne dégrade l’ADN que dans le sens 5' → 3'.
a. Cette excision s’effectue à partir d’une discontinuité préexistante du brin néosynthétisé, telle qu’une extrémité de fragments d’Okazaki ou une entaille (nick) de réplication, et progresse jusqu’au site du mésappariement après l’incision supplémentaire introduite par l’endonucléase PMS2.
- Si la coupure se trouve du côté 5' du mismatch, dans le cas d'une extrémité d’un fragment d’Okazaki ou une entaille laissée par la réplication, l’exonucléase EXO1 dégrade directement l’ADN de 5' → 3' en direction du mésappariement.
- Si la coupure se situe du côté 3' du mismatch, le système MMR introduit généralement une incision supplémentaire par MutLα (PMS2) du côté 5' du mismatch.
Cette nouvelle coupure permet alors à EXO1, qui agit uniquement 5' → 3', de dégrader l’ADN jusqu’au site du mésappariement.
b. L’excision élimine typiquement un segment de plusieurs dizaines à plusieurs centaines de nucléotides, garantissant l’élimination complète du mésappariement.
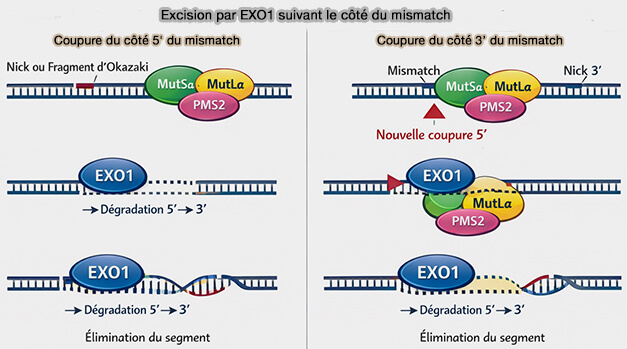
(Figure : vetopsy.fr)
2. La progression de l’excision est contrôlée par les complexes MutS et MutL qui restent associés à l’ADN et coordonnent l’extension de la région simple brin générée par l’excision, en stabilisant l’ADN simple brin et en orientant l’activité d’EXO1 vers le site du mésappariement.
Cette organisation empêche l’excision du brin matrice et limite la propagation de la dégradation au-delà de la zone nécessaire.
3. Une fois le segment erroné éliminé, la région simple brin est resynthétisée en utilisant le brin parental intact comme matrice.
L’ADN polymérase δ restaure la séquence correcte en rétablissant l’appariement canonique des bases, cette synthèse étant couplée à la progression de la pince PCNA le long de l’ADN.
4. La brèche résiduelle est ensuite scellée par une ADN ligase, principalement l’ADN ligase I (LIG1) dans le contexte de la réplication, ce qui rétablit la continuité du squelette phosphodiester du brin réparé.
Importance biologique et pathologique du système MMR
1. Une défaillance du système MMR entraîne une accumulation rapide de mutations ponctuelles et une instabilité des microsatellites (MSI), caractérisée par des variations de longueur des séquences répétées (Microsatellite instability and immune checkpoint inhibitors: toward precision medicine against gastrointestinal and hepatobiliary cancers 2019).
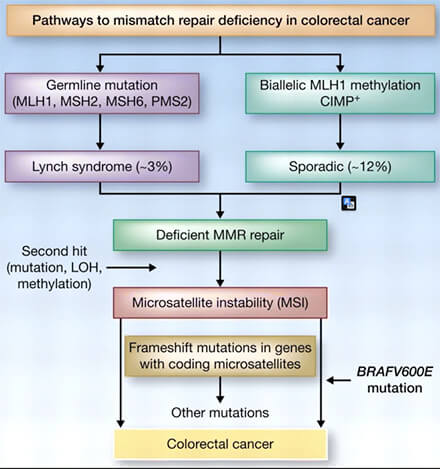
(Figure : vetopsy.fr d'après Pastor et Schlom)
Chez l’humain, ces altérations sont associées au syndrome de Lynch, une prédisposition héréditaire aux cancers colorectaux et à d’autres tumeurs, liée à des mutations dans les gènes MMR tels que MLH1, MSH2, MSH6 ou PMS2 (Immunology of Lynch Syndrome 2021).
2. Au-delà de la correction des erreurs de réplication, le système MMR intervient également :
- lors de la recombinaison homologue (HR), en détectant les mésappariements présents dans les hétéroduplex formés lors de l’appariement entre séquences d’ADN imparfaitement homologues (
 recombinaison homologue ou HR).
recombinaison homologue ou HR). - lors de certaines réponses cellulaires aux dommages de l’ADN, notamment en détectant des bases altérées ou des mésappariements persistants qui peuvent déclencher des voies de signalisation conduisant à l’arrêt du cycle cellulaire ou à l’apoptose, en particulier lorsque les lésions ne peuvent pas être correctement réparées.
Réparation des cassures double brin
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation