Le domaine AAA ou AAA+ est un domaine ATPasique spécifique de la famille des protéines AAA (ATPases Associated with diverse cellular Activities), le plus souvent sous forme hexamérique.
Le domaine AAA est quelquefois appelé aussi AAA+.
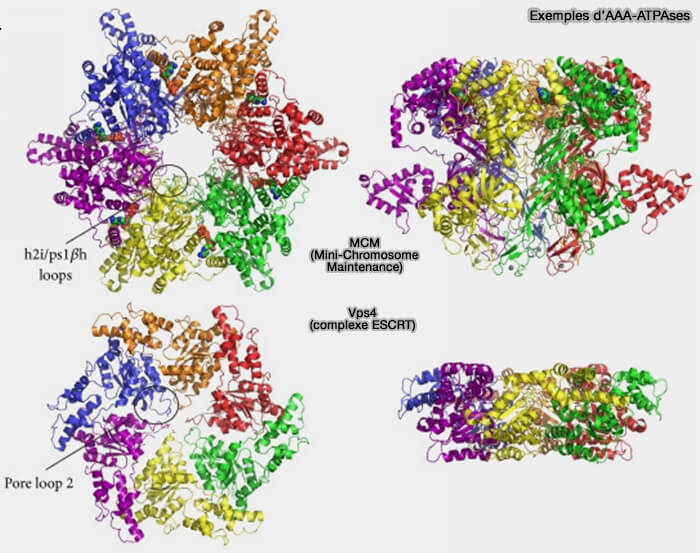
Exemples de protéines AAA ou AAA-ATPases
(Figure : vetopsy.fr d'après Miller et Enemark)
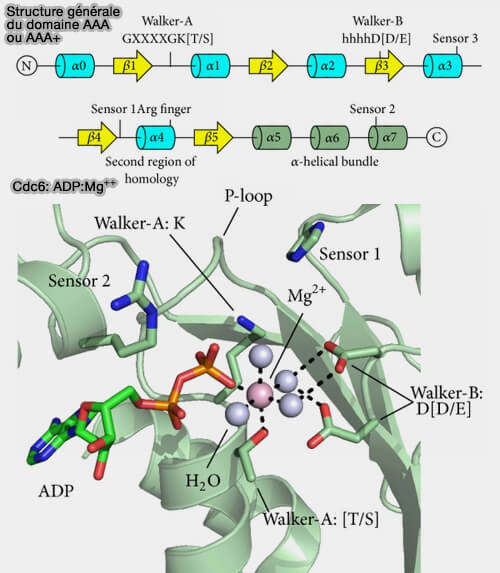
Structure générale du domaine AAA
(Figure : vetopsy.fr d'après Miller et Enemark)
Remarque : la structure en sandwich α-β-α est l’un des plus anciens, sinon le plus ancien repliement protéique connu, qui se décline en deux versions, P-loop et pli de Rossmann.
2. Les caractéristiques qui distinguent les membres de la famille AAA+ des autres NTPases à P-loop comprennent :
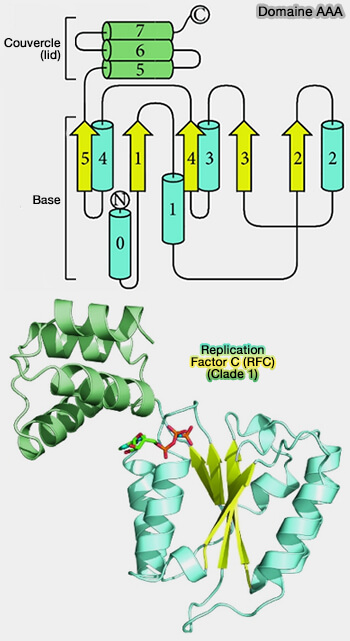
Structure du domaine AAA et clade 1
(Figure : vetopsy.fr d'après Miller et Enemark)
l’insertion de β4 entre β1 et β3,
l’absence d’un brin β antiparallèle adjacent à β5,
l’absence de tout brin supplémentaire directement adjacent à β5 ou β2.
3. Le site ATPase AAA+ se trouve à l’interface des sous-unités adjacentes d’un complexe protéique.
Motifs de l'activité ATPase
Au sein de ce domaine, on retrouve les motifs structuraux essentiels à l'activité ATPase.
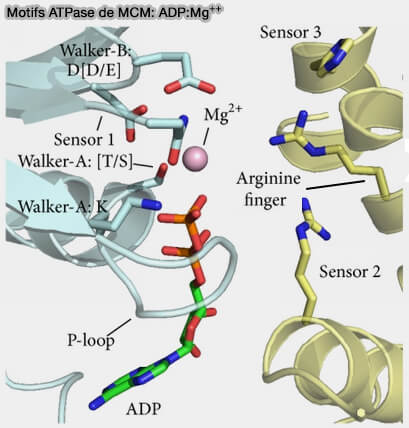
1. Le motif A de Walker est impliqué dans la liaison de l'ATP, i.e. GxxxxGK[T/S], et forme une boucle entre β1 et α1, qui correspond à la boucle P (Phosphate-binding loop) canonique qui est aussi présente dans les GTPases ( domaine G).
Elle interagit directement avec le groupe β-phosphate, mais pas avec le ribose comme dans le pli de Rossmann.
Elle stabilise la charge négative des phosphates grâce au Mg++, quelquefois avec le Ca++ dans d'autres NTPases, alors que le pli de Rossmann canonique n'en possède pas, contrairement à celui de la tubuline.
Elle positionne l’ATP dans la bonne orientation pour que l’hydrolyse puisse se produire efficacement.
Elle détecte l'état de liaison selon que l’ATP, l’ADP ou aucun nucléotide est lié, ce qui influence la conformation globale de la protéine.
2. Le motif B de Walker B, nécessaire à l'hydrolyse de l'ATP, i.e. hhhhD[D/E] dans lequel h représente un résidu hydrophobe, s’étend sur β3.
Motifs ATPase de MCM: ADP:Mg++
(Figure : vetopsy.fr d'après Miller et Enemark)
3. Le motif SRH (Second Region of Homology), motif C-terminal du motif Walker B, i.e. 15 à 20 résidus qui s'étend sur β4, l'hélice α4 et la boucle α4/β5 et comprend le senseur 1 et le motif en doigt (finger) d'arginine nécessaires à l'hydrolyse de l'ATP.
Ils coordonnent l’hydrolyse des nucléotides et propagent les changements de conformation associés à l’hydrolyse des nucléotides entre les sous-unités des complexes protéiques AAA+.
Ce domaine, caractéristique des AAA+, n'existe pas dans les autres NTPases.
Il fonctionne de concert avec le glutamate Walker-B pour orienter correctement la molécule d’eau nucléophile qui subit une attaque sur le γ-phosphate de la molécule ATP liée.
Les résidus Walker A/B et le senseur 1 du site ATPase sont tous situés sur la même sous-unité, d'où le terme de cis-agissant, i.e. cis-acting.
b. Le doigt d’arginine, situé dans la boucle entre α4 et β5, est dérivé de la sous-unité voisine, d'où le terme de trans-agissant.
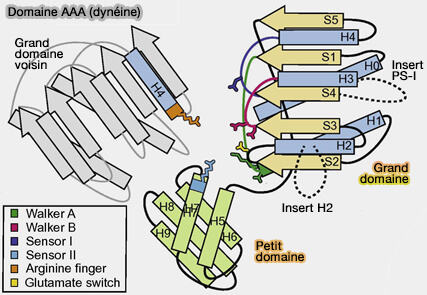
Domaine AAA de la dynéine
(Figure : vetopsy.fr d'après Gleave et coll)
4. Le motif senseur 2, constitué d’arginine ou de lysine et situé près au début de l'hélice α7, est impliqué dans les changements conformationnels associés à un cycle de liaison et d’hydrolyse de l’ATP.
a. Ce résidu interagit avec l'α-phosphate de la molécule d’ATP liée, sa mutation diminue la liaison des nucléotides.
Les membres de la famille AAA, par opposition aux membres de la famille AAA+, possèdent généralement un résidu d’alanine dans cette position.
b. Le senseur 2 fonctionne comme un résidu cis-agissant dans les protéines AAA+ qui contiennent des couvercles C-terminaux et est un résidu trans-agissant dans les protéines dépourvues d’un domaine de paupière canonique α-hélicoïdal.
Un exemple de senseur 2 en action est dans les protéines MCM, où une insertion dans le faisceau hélicoïdal α C-terminal perturbe le domaine canonique du couvercle et positionne l’arginine du senseur 2 comme un résidu trans-agissant (Analysis of the crystal structure of an active MCM hexamer 2014).
Cette région contient généralement une glycine conservée, ou un résidu tout aussi petit, qui forme une coiffe à l’extrémité N-terminale, suivie d’un autre résidu conservé par la famille, i.e. autre glycine ou un résidu hydrophobe.
Le N-linker est la région N-terminale conservée à cette séquence de dipeptides qui s’étend perpendiculairement aux brins β du noyau α-β-α.
Il contribue à la fois à la poche de liaison à l’ATP et sert à connecter le domaine AAA+ à d’autres domaines au sein d’une protéine.





 domaine G).
domaine G).