La phosphorylation, l’ubiquitination, la SUMOylation, l’ADP-ribosylation et la citrullination sont des modifications post-traductionnelles des histones qui modulent la structure de la chromatine et participent à la régulation transcriptionnelle, au cycle cellulaire et à la réponse aux dommages de l’ADN.
1. La phosphorylation des histones est catalysée par différentes kinases nucléaires, appelées writers dans le cadre du code des histones, qui sont souvent impliquées dans les voies de signalisation ou dans le cycle cellulaire.
La kinase DNA-PK (DNA-dependent protein kinase) peut également phosphoryler H2AX sur la sérine 139 et contribue ainsi à la réponse cellulaire aux dommages de l’ADN.
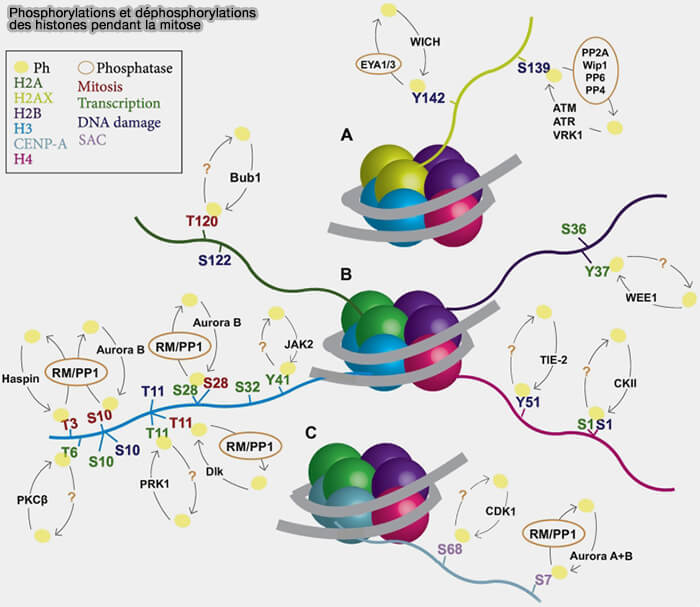
Phosphorylations et déphosphorylations des histones pendant la mitose
(Figure : vetopsy.fr d'après Gil et Vagnarelli)
La phosphorylation introduit une charge négative supplémentaire sur les histones, ce qui modifie les interactions entre les histones et l’ADN ou entre nucléosomes et participe à des transitions rapides entre états condensés (hétérochromatine) et accessibles de la chromatine (euchromatine).
Cette modification peut entraîner plusieurs effets structuraux :
déstabilisation locale des interactions histone-ADN, facilitant l’accès aux facteurs de transcription.
modification de l’organisation nucléosomique lors de réponses cellulaires rapides, comme la réparation de l’ADN.
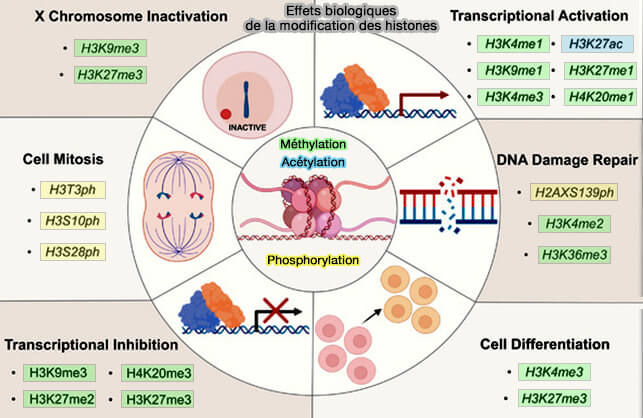
Effets biologiques principaux de la modification des histones
(Figure : vetopsy.fr d'après Liu et coll)
Reconnaissance par des protéines effectrices
Contrairement aux marques d’acétylation ou de méthylation, les lecteurs spécialisés des histones phosphorylées sont relativement peu nombreux.
La phosphorylation agit souvent comme un signal transitoire de signalisation cellulaire, notamment dans la mitose ou la réponse aux dommages de l’ADN (DDR), et sa reconnaissance dépend fréquemment de contextes protéiques plus larges plutôt que de domaines de lecture strictement dédiés.
1. Les histones phosphorylées peuvent être reconnues par des protéines spécialisées, appelées readers, qui possèdent des domaines capables de reconnaître spécifiquement les résidus phosphorylés.
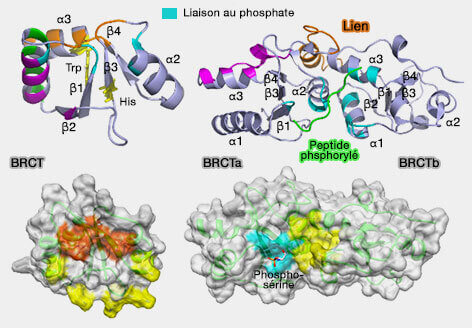
BRCT simple et en tandem
(Figure : vetopsy.fr d'après Sheng et coll)
Ces domaines reconnaissent spécifiquement des motifs peptidiques contenant des sérines ou thréonines phosphorylées, souvent dans un contexte de signalisation déclenché par les kinases ATM, ATR ou DNA-PK. Dans la chromatine, les domaines BRCT permettent notamment la reconnaissance indirecte de la marque γH2AX (H2AXS139ph) via des protéines comme MDC1 (Mediator of DNA Damage Checkpoint 1), dont les domaines BRCT se lient spécifiquement à cette histone phosphorylée, ce qui contribue au recrutement de nombreux facteurs de réparation autour des cassures double brin de l’ADN.
b. Les domaines FHA (Forkhead-associated domains) constituent une autre famille de modules de reconnaissance des phospho-résidus.
Ces domaines reconnaissent préférentiellement des thréonines phosphorylées (pThr) dans des motifs spécifiques de protéines cibles.
Les protéines contenant des domaines FHA interviennent fréquemment dans les voies de signalisation et les checkpoints du cycle cellulaire, ainsi que dans la réponse aux dommages de l’ADN.
Leur interaction avec des protéines phosphorylées contribue à organiser des complexes multiprotéiques impliqués dans la réparation de l’ADN et la régulation de la chromatine.
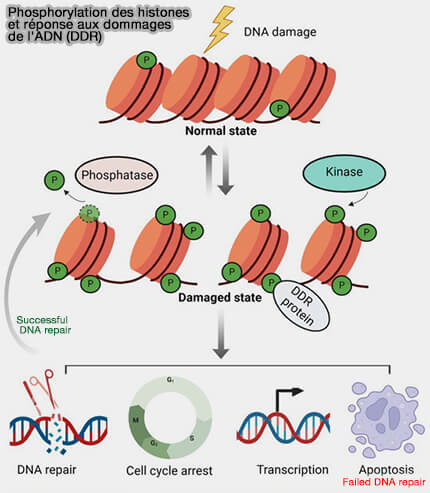
Phosphorylation des histones et dommages de l'ADN
(Figure : vetopsy.fr d'après Gong et coll)
c. Les protéines 14-3-3 possèdent des domaines capables de reconnaître des motifs sérine/thréonine phosphorylés.
Elles peuvent se lier à certaines histones phosphorylées, par exemple H3S10ph ou H3S28ph, et participent alors à des processus de régulation transcriptionnelle dépendants de la phosphorylation des histones, notamment lors de l’activation transcriptionnelle induite par des voies de signalisation.
2. Ces interactions permettent le recrutement de complexes protéiques impliqués dans différentes fonctions nucléaires comme :
la réparation de l’ADN,
la régulation transcriptionnelle,
l'organisation du chromosome mitotique.
Remarque : la phosphorylation des histones peut agir en combinaison avec d’autres modifications post-traductionnelles, notamment l’acétylation.
Par exemple, la phosphorylation de H3S10 peut favoriser l’acétylation du résidu voisin H3K14, formant un module de phospho-acétylation associé à l’activation transcriptionnelle (enhancer) de certains gènes.
a. H2AX est une variante de l’histone H2A incorporée dans les nucléosomes, caractérisée par une extension C-terminale spécifique contenant le motif SQ(E/D), permettant sa phosphorylation sur la sérine 139.
Les kinases ATM, ATR et DNA-PKcs phosphorylent la sérine 139 de la variante histonique H2AX, pour former la marque γH2AX qui signale la présence de cassures double brin de l’ADN.
ATM agit principalement en réponse aux cassures double brin détectées par le complexe MRN,
ATR intervient surtout lors de stress réplicatif associé à la formation d’ADN simple brin recouvert de RPA,
DNA-PKcs est recrutée directement aux extrémités des cassures double brin par le complexe Ku70-Ku80 dans la voie NHEJ.
b. Cette modification apparaît rapidement autour des sites de dommages et forme des foyers nucléaires détectables par immunofluorescence, permettant de visualiser et de quantifier les dommages à l’ADN dans les cellules exposées à des radiations ionisantes, à des agents génotoxiques ou lors d’instabilités génomiques.
Elle est ainsi fréquemment utilisée comme indicateur expérimental de stress génotoxique et d’instabilité génomique dans les études sur le cancer et le vieillissement.
3. Les phosphorylations majeures sont résumées dans le tableau suivant.
3. Des altérations de la phosphorylation des histones peuvent être associées à différentes pathologies.
Par exemple, des anomalies dans la phosphorylation de H3S10 ou dans la signalisation γH2AX sont observées dans certains cancers et peuvent refléter une dérégulation de la transcription, du cycle cellulaire ou de la réponse aux dommages de l’ADN.
Au cours du vieillissement, des modifications dans la phosphorylation de H2A.X, H3 ou H1 peuvent également altérer l’efficacité de la réponse aux dommages de l’ADN et contribuer à l’accumulation d’instabilités génomiques.
De manière plus générale, des défauts dans les voies de phosphorylation impliquées dans la réponse au stress cellulaire ou dans la mitose peuvent participer à des processus pathologiques, notamment dans certaines maladies génétiques ou neurodégénératives.
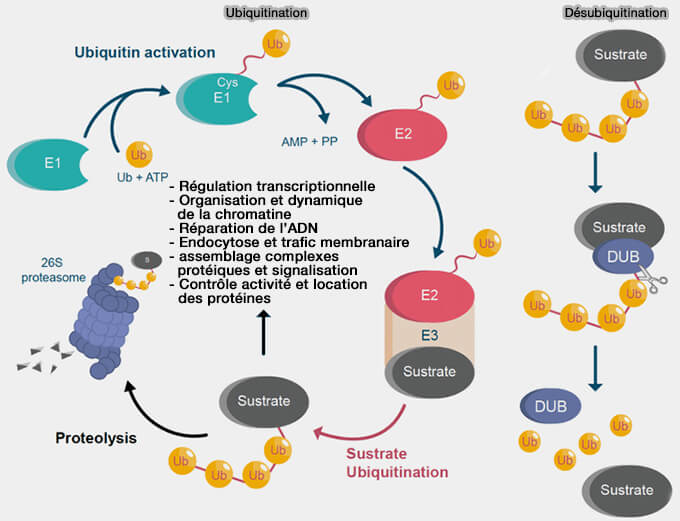
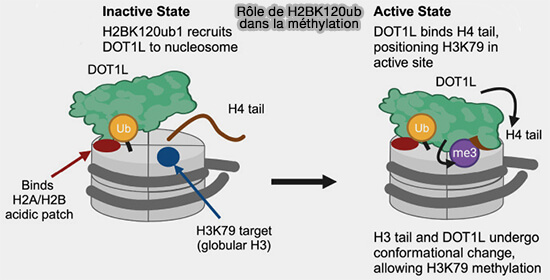
1. H2AK119ub est la marque d’ubiquitination des histones la plus étudiée.
Elle est associée à la répression transcriptionnelle, notamment via le complexe PRC1 (Polycomb Repressive Complex 1), à ne pas confondre avec PRC1 (Protein Regulator of Cytokinesis 1), une protéine associée aux microtubules impliquée dans l’organisation du fuseau mitotique (MAP organisatrice).
Cette modification est associée à une répression transcriptionnelle durable et participe à l’organisation de l’hétérochromatine facultative, en lien fonctionnel avec l’activité du complexe PRC2 qui dépose la marque H3K27me3.
Lors du développement et de la différenciation cellulaire, le complexe PRC1 ubiquitine ainsi H2AK119 afin de maintenir silencieux certains gènes de développement.
Dans ce contexte, l’ubiquitination de certaines histones peut faciliter le recrutement de protéines impliquées dans la réparation de l’ADN et contribuer à la réorganisation locale de la chromatine autour des sites de lésions.
Remarque : les histones ubiquitinées peuvent être reconnues par des protéines spécialisées, appelées readers, qui possèdent des domaines UBD (Ubiquitin-Binding Domains), capables de reconnaître l’ubiquitine ou les nucléosomes ubiquitinés pour participer au recrutement de complexes de réparation ou de régulation transcriptionnelle.
4. Des altérations des voies d’ubiquitination des histones peuvent être observées dans certaines pathologies.
Par exemple, des dérégulations des marques H2AK119ub ou H2BK120ub peuvent perturber l’équilibre entre activation et répression transcriptionnelle dans certains cancers.
Des mutations affectant certaines ligases d’ubiquitine impliquées dans ces processus peuvent également contribuer à des anomalies du développement ou à des altérations de l’organisation de la chromatine.
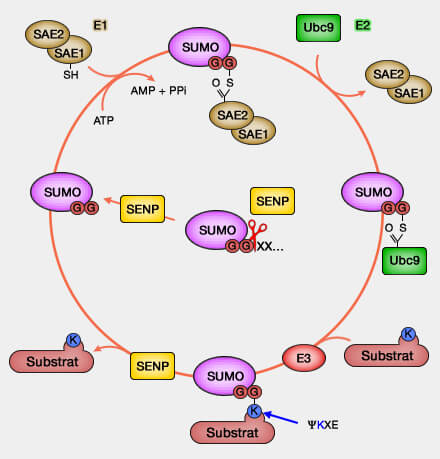
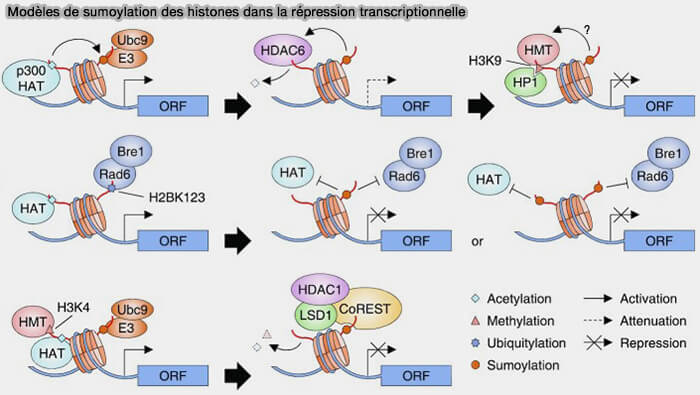
Remarque : les histones sumoylées peuvent être reconnues par les domaines SIM (SUMO Interaction Motifs) qui permettent le recrutement de complexes répressifs ou de régulateurs transcriptionnels.
3. Des altérations de la régulation de la sumoylation peuvent être observées dans certaines pathologies, notamment dans certains cancers ou lors du vieillissement, où elles peuvent contribuer à des perturbations de la régulation transcriptionnelle et de l’organisation de la chromatine.
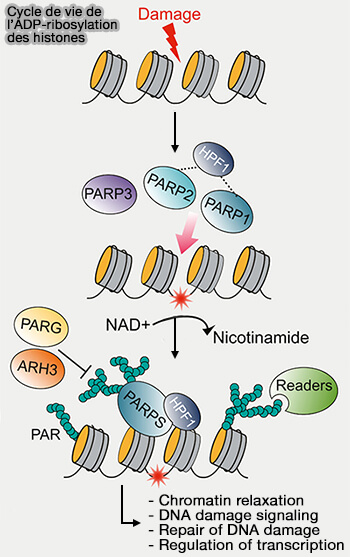
ADP-ribosylation des histones
1. L’ADP-ribosylation consiste en l’ajout d’un ou de plusieurs groupements ADP-ribose sur certains résidus des histones, par transfert d’unités ADP-ribose issues du NAD+ ( rôles de NAD+).
ADP-rybosylation des histones
(Figure : vetopsy.fr d'après Özdemir et coll)
a. Cette modification est catalysée principalement par des enzymes de la famille des PARP (Poly-ADP-ribose polymerases) ou ARTD (EC 2.4.2.30) qui utilisent le NAD+ comme substrat et transfèrent une unité ADP-ribose sur les protéines cibles en libérant une molécule de nicotinamide.
L’ADP-ribosylation peut être mono-ADP-ribosylation ou poly-ADP-ribosylation, selon le nombre d’unités ajoutées.
b. La modification est réversible grâce à des enzymes hydrolysant l’ADP-ribose, notamment la poly-ADP-ribose glycohydrolase PARG (EC 3.2.1.143) et certaines ADP-ribosylhydrolases telles que ARH3 (EC 3.2.1.191).
Remarque : les histones poly-ADP-ribosylées peuvent être reconnues par :
les macrodomains, domaine protéique d’environ 130–190 acides aminés,
a. L’activation rapide des poly(ADP-ribose) polymérases (PARP), en particulier de PARP1, permet leur fixation sur l’ADN endommagé grâce à ses domaines de liaison à l’ADN de type zinc finger (Zn1 et Zn2) capables de reconnaître les discontinuités du squelette phosphodiester au niveau des cassures.
b. Cette activation déclenche un changement conformationnel activant le domaine catalytique de l’enzyme et entraîne la synthèse locale de chaînes de poly(ADP-ribose) à partir du NAD+.
PARP1 catalyse alors son auto-ADP-ribosylation, qui entraîne la formation de chaînes de poly(ADP-ribose) fortement chargées négativement diminuant l’affinité de l’enzyme pour l’ADN et favorisant sa dissociation du site de cassure, tout en créant une plateforme de liaison permettant le recrutement de nombreuses protéines impliquées dans la réponse aux dommages de l’ADN ( réparation de l'ADN).
c. La protéine HPF1 (Histone PARylation Factor 1) peut s’associer au complexe PARP1 fixé à l’ADN endommagé et former avec lui un site catalytique composite qui modifie la spécificité enzymatique de PARP1 et oriente l’ADP-ribosylation vers les histones de la chromatine environnante, notamment sur des résidus sérine des histones H3 et H2B.
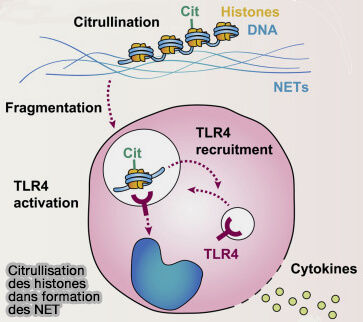
1. Cette réaction est catalysée par les PAD (peptidyl-arginine déiminases), notamment PAD4 qui est la principale enzyme nucléaire (EC 3.5.3.15).
La citrullination entraîne la perte d’une charge positive de l’arginine, ce qui diminue les interactions entre les histones et l’ADN et favorise ainsi la décondensation de la chromatine.
Contrairement à la plupart des autres modifications des histones, la citrullination ne possède pas d’enzyme de
décitrullination connue et est généralement éliminée par le renouvellement ou la dégradation des histones.
2. La citrullination intervient notamment dans certains processus de régulation transcriptionnelle et dans des contextes cellulaires particuliers.
Un exemple bien connu est la formation des NETs (Neutrophil Extracellular Traps) lors de la NETosis des neutrophiles ( NET dans l'athérosclérose).
La citrullination massive des histones provoque une forte décondensation de la chromatine, permettant aux neutrophiles de libérer leur ADN dans le milieu extracellulaire.
Les histones citrullinées associées à l’ADN extracellulaire contribuent ensuite à l’activation du récepteur de l’immunité TLR4 (Toll-like receptor 4) sur des cellules cibles comme les macrophages ou les cellules dendritiques.
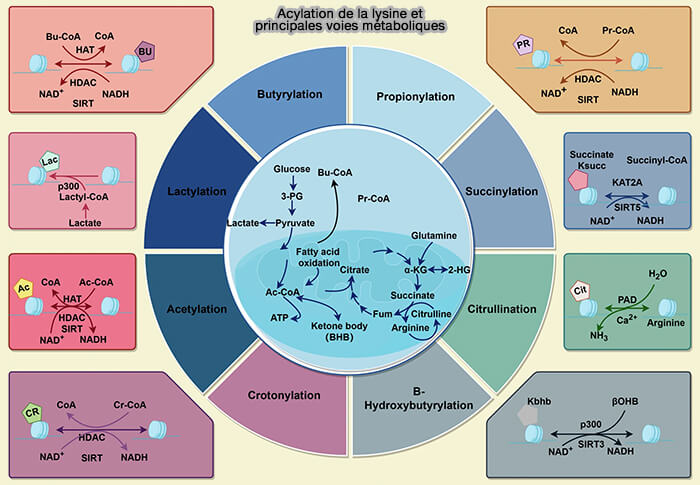
En plus des modifications classiques des histones, de nombreuses nouvelles modifications post-traductionnelles des histones (HPTM) ont été identifiées ces dernières années grâce aux progrès de la protéomique et de la spectrométrie de masse
2. Ces modifications sont souvent liées au métabolisme cellulaire et pourraient contribuer à l’intégration des signaux métaboliques dans la régulation de la chromatine et de l’expression génique.Acylation de la lysine et principales voies métaboliques
(Figure : vetopsy.fr d'après Yao et coll)




 dommages de l'ADN).
dommages de l'ADN).